

gistologia
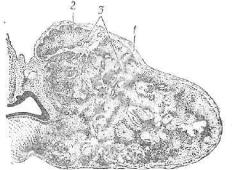
.pdfРис. 221. Гонадотропоцит передней доли аденогипофиза:
1 — ядро; 2 — комплекс Гольджи*. з — секреторные гранулы; 4 — накопитель ные гранулы; 5 — митохондрии; в — цистерны гранулярной эндоплазматиче ской сети.
терстициальных клеток в семенниках (рис. 221). В центральной зоне гонадотропного базофила находится макула. Это расширен1- ная полость комплекса Гольджи, оттесняющая ядро," многочислен ные мелкие митохондрии, мембраны эндоплазматической сети на. периферию клетки. Базофильные гонадотропоциты содержав зер нистость, равную около 200—300 нм в диаметре.
При недостаточности половых гормонов в организме диаметр зернистости увеличивается. После кастрации животных базофиль ные гонадотропоциты превращаются в клетки кастрации: крупная вакуоль занимает всю центральную часть клетки. Последняя при обретает кольцевидную форму.
Тиреотропные базофилы (рис. 222)','— угловатые клетки с мел1- кой (80—150 нм) заполняющей всю цитоплазму зернистостью-, Если
Рис. 222. Тиреотропоцпт передней доли аденогипофиза (электронная микрофотография):
1 — ядро; 2 — секреторные гранулы; 8 — соматотропоцит (по Долан и Сёлоши),
организм испытывает недостаток гормонов щитовидной железы, то» развиваются клетки тиреодэктомии. Они увеличены в размерег с расширенными цистернами эндоплазматической сети, поэтому цитоплазма имеет ячеистый вид, более крупные гранулы секрета,
Хромофобные клетки составляют 60—70% всех клеток перед ней части гипофиза. Это сборная группа, так как в ее состав вхо дят разные по значению клетки: камбиальные, клетки на разной* стадии дифференциации; не накопившие еще специфическую* зернистость; клетки, выделившие секрет. Из камбиальных клеток в дальнейшем развиваются ацидофильные и базофильные аденоциты.
Промежуточная часть аденогипофиза представлена нескольки1- ми рядами слабобазофильных клеток. Продуцируемый аденоцита-
304 |
20 Заказ Ж 908 |
30 |
$ |
|
ми секрет накапливается в промежутках между клетками, что •способствует образованию фолликулоподобных структур. Клетки промежуточной части аденогипофиза полигональной формы, со-/ держат мелкие гликопротеидные гранулы размером 200—300 нм/ В промежуточной зоне синтезируются меланотропин, регулирую/ щий пигментный обмен, и липотропин — стимулятор жирового обмена.
Туберальная часть аденогипофиза по |
своей структуре |
сходна |
с промежуточной частью. Она прилегает |
к гипофизарпой |
ножке |
и медиальной эминенции. Клетки этой зоны характеризуются сла бой базофилией и трабекулярным расположением. Функция туберальной части окончательно не выяснена.
Выше говорилось о том, что гормонообразовательная функция аденогипофиза регулируется гипоталамусом, с которым он обра зует единую гипоталамоаденогипофизарную систему. Морфофункционально эта связь проявляется в следующем: верхняя гипофизарная артерия в медиальной эминенции образует первичную ка пиллярную сеть. Аксоны мелких нейросекреторных клеток ядер медиобазального гипоталамуса на сосудах первичной капиллярной сети образуют аксоваскулярные синапсы. Нейрогормоны, проду цируемые этими нейросекреторными клетками, по их аксонам пе ремещаются в медиальную эминенцию. Здесь они накапливаются, а затем через аксоваскулярные синапсы поступают в капилляры первичной сосудистой сети. Последние собираются в портальные вены, которые направляются вдоль гипофизарной ножки в аденогипофиз. Здесь вновь они распадаются и образуют вторичную ка пиллярную сеть. Синусоидные капилляры этой сети оплетают трабекулы секретирующих аденоцитов.
Кровь, оттекающая по венам от вторичной сосудистой сети, со держит аденогипофизарные гормоны, которые через общий крово ток, то есть гуморальным способом, регулируют функции эндо кринных желез периферического звена.
Нейрогипофиз (задняя доля) развивается из мозгового карма на, поэтому он построен из нейроглии. Его клетки — питуициты веретенообразной или отростчатой формы. Отростки питуицитов контактируют с кровеносными сосудами. В заднюю долю входят крупные пучки нервных волокон, образованные аксонами нейро секреторных клеток паравентрикулярных и супраоптических ядер передней зоны гипоталамуса. Нейросекрет, образовавшийся этими клетками, перемещается вдоль аксонов в нейрогипофиз в виде сек реторных капель. Здесь они оседают в виде накопительных телец, или терминалей, которые контактируют с капиллярами.
Следовательно, гормоны нейрогипофиза — окситоцин и вазопрессин синтезируются не структурами нейрогипофиза, а в пара вентрикулярных и супраоптических ядрах. Затем, как говорилось выше, по нервным волокнам гормоны поступают в нейрогипофиз, где они накапливаются и откуда поступают в кровяное русло. По этому нейрогипофиз и гипоталамус тесно связаны и формируют единую гипоталамонейрогипофизарную систему.
306
Окситоцин стимулирует функцию гладких мышц матки, этим способствует выделению секрета маточных желез; при родах вы зывает сильное сокращение мышечной оболочки стенки матки; ре гулирует сокращение мышечных элементов молочной железы.
Вазопрессин суживает просвет кровеносных сосудов и повы шает кровяное давление; регулирует водный обмен, так как влияет на обратное всасывание (реабсорбцию) воды в канальцах почки-
ЭПИФИЗ
Эпифиз (верхний мозговой придаток), или шишковидное тело, раз вивается в виде непарного утолщения задней части крыши проме жуточного мозга. В закладке эпифиза принимает участие невральная эктодерма, дающая начало клеткам двух типов паренхимы^ органа: секреторным клеткам — пипеалоцитам и нейроглиальным клеткам — глиоцитам. Из мезенхимы развивается строма органа: соединительнотканные капсула, трабекулы и неполные перего родки.
Эпифиз функционирует только у молодых животных. В даль нейшем он подвергается инволюции.
Расположен эпифиз между полушариями большого мозга w мозжечком. Снаружи он окружен мягкой мозговой оболочкой и соединительнотканной капсулой, от которой во внутрь органа отхо дят тонкие трабекулы и неполные перегородки, делящие орган на дольки (рис. 223). Паренхима долек построена из шшеалоцитовг глиоцитов, встречаются также лимфоциты, тканевые базофплы. пигментные клетки и мозговой песок.
П и н е а л о ц и т ы характеризуются складчатым ядром с хорошо выраженными ядрышками и многочисленными длинными отро стками (рис. 224). Последние направляются к соединительноткан ным перегородкам, где заканчиваются булавовидными расшире ниями вблизи кровеносных сосудов или на их стенках. В цитоплаз ме найдены одиночные свободные рибосомы, короткие профили гранулярной эндоплазматической сети, митохондрии, комплекс Гольджи, центриоли; интенсивно развиты микротрубочки; встре чаются включения липидов и липохрома. Среди пинеалоцитов раз личают две разновидности:
главные и темные клетки. Первые характеризуются однородной цитоплазмой, вторые — меньшие по раз меру — имеют в цитоплаз ме ацидофильную или базофильную зернистость.
Рис. 223. Эпифиз:
1 — капсула; 2 — паренхима дольки; 3 — трабекулы (по Шафферу).
20* |
ЗОГ |

Г л ы о ц и т ы — это отростчатые клетки с базофильной цито плазмой и вытянутым ядром, хроматин которого находится в более конденсированном состоянии (рис. 225). Цитоплазма отличается множеством микрофиламентов диаметром 5—б нм. Органеллы развиты слабее, чем в пинеалоцитах.
Пинеалоциты секретируют серотонин, образующий мелатонин — гормон-антагонист меланоцитсинтезирующего гормона пе редней части аденогипофиза. Синтез мелатонина находится в пря мой зависимости от воздействия светового фактора на организм. При недостаточном световом раздражении этот процесс усили вается.
Кроме того, серотонин и мелатонин угнетают функцию органов размножения, оказывают воздействия на другие эндокринные же лезы, часто как антагонисты.
Пинеалоциты вырабатывают и гормоны белковой природы. К ним относят шгаеальный антигонадотропин. Действуя на аденогипофиз, он одновременно с гипоталамусом ослабляет синтез гонадотропных гормонов, снижая половую активность организма.
При инволюции эпифиз прорастает соединительной тканью, образуется мозговой песок — слоистые, округлые известковые от ложения, состоящие из фосфата и карбоната со следами магния.
ПЕРИФЕРИЧЕСКИЕ ЗВЕНЬЯ ЭНДОКРИННОЙ СИСТЕМЫ
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа образуется из энтодермального эпителия не парного срединного выроста вентральной стенки передней кишки. Эпителиальные клетки формируют сложную систему тяжей, из мезенхимы развивается соединительная ткань, которая покрывает зачаток снаружи и врастает в него. Из материала непарного эм брионального органа образуются две доли, соединенные перешей ком. Последний сохраняется на всю жизнь только у крупного ро гатого скота и свиней.
Щитовидная железа расположена в области шеи по обеим сто ронам трахеи, позади щитовидного хряща.
Снаружи щитовидная железа покрыта соединительнотканной капсулой, от которой в глубь органа отходят перегородки, разде ляющие паренхиму органа на дольки, а дольки на замкнутые пу зырьки — фолликулы (рис. 226).
Основной морфофункциональной структурой щитовидной же лезы является фолликул — замкнутый округлый или овальный пузырек. Размеры фолликулов варьируют от 0,02 до 0,9 мм в диа метре. В фолликуле различают стенку и полость, заполненную коллоидом. Стенка фолликула состоит из однослойного эпителия, расположенного на базальной мембране.
309
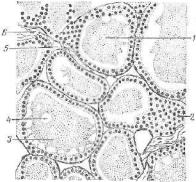
Рис. 226. Щитовидная железа лоша-j ди:
1 — фолликул; 2 — стенка фолликула*. з — коллоид; 4 — вакуоль; 5 — напил* ляр; в —• соединительная ткань.
Форма клеток обусловлена функциональной активностью щитовидной железы и может быть или плоской, или кубичес кой, или столбчатой (цилиндри ческой) . Если железа характери зуется умеренной функцией, токлетки фолликула имеют куби
ческую форму. При повышенной деятельности железы (гиперфунк ция) отмечают усиленное поступление гормона в кровь, клетки приобретают столбчатую форму (см. цв. табл. VII—Б). Снижение функциональной активности железы (гипофункция) сопряжено с увеличением диаметра фолликулов и накоплением коллоида в их полостях. Резко уменьшается при этом высота клеток. Они стано вятся уплощенными (В).
Функциональное состояние железы влияет и на консистенцию коллоида. При умеренной функции коллоид гомогенен и заполняет всю полость фолликула. При гиперфункции коллоид более жидкой консистенции, имеет пенистый вид, много вакуолей; снижается содержание коллоида в фолликулах. При гипофункции коллоид; сгущается ж уплотняется.
Внутренняя выстилка фолликулов представлена клетками двух типов: фолликулярными клетками (тироцитами) и околофоллпкулярными клетками (К-клетками). Последние встречаются реже и могут располагаться не только в стенке фолликула, но и между ними. Функция тироцитов сводится к синтезу йодсодержащих гор монов тироксина и трийодтиронина. Они регулируют окислитель ные процессы, влияющие на все виды, обмена веществ, протекаю щие в организме. Гормонообразовательная функция фолликуляр ных клеток стимулируется тиротропными гормонами, поэтому они относятся к той группе эндокринных клеток, функция которых зависит от передней доли гипофиза.
Околофолликулярные клетки вырабатывают йоднесодержащий гормон — кальцитонин (тирокальцитонин), который снижает со держание кальция в крови и является антагонистом паратгормона, синтезирующего околощитовидной железой. Гормональна» функция околофолликулярных клеток (К-клеток) не зависит от передней доли гипофиза.
Фолликулярные клетки имеют светлое, центрально располо женное округлое ядро. В цитоплазме базального полюса находятся хорошо развитые мембранные структуры гранулярной эндоплазматической сети, митохондрии с незначительным числом криет,.
ЗЮШ^Н ^КЩ1 |
1 |
Плазмолемма образует базальную складчатость. Над ядром или около него лежит комплекс Гольджи, лизосомы. В цитоплазме име ются мелкие капли коллоида. Плазмолемма апикального полюса образует микроворсинки, увеличивающие поверхность контакта тироцитов с полостью фолликула. Между собой клетки соединя ются при помощи пятен слипания и терминальных пластинок.
Околофолликулярные (светлые) клетки — К-клетки расположе ны в стенке фолликулов или в составе межфолликулярных остров ков, лежащих в межфолликулярной соединительной ткапи. Это светлые, крупные, овальные клетки, апикальная поверхность ко торых не контактирует с полостью и коллоидом фолликула. В К-клетках хорошо развиты гранулярная эндоплазматическая сеть, комплекс Гольджи, что свидетельствует об интенсивном сиптезе белков; цитоплазма содержит белковые секреторные гранулы 0,1—0,4 мкм в диаметре, незначительное количество митохондрий. Особенностью этих клеток является неспособность поглощать йод.
Составными клетками межфолликулярных островков также яв ляются эпителиальные клетки, которые выполняют функцию ис точника развития новых фолликулов.
Снаружи фолликулы покрыты базальной мембраной. Фолли кулы отграничены тонкими прослойками рыхлой соединительной ткани, интенсивно снабженной гемо- и лимфососудистой сетью. Межфолликулярная соединительная ткань, соединяясь с междольковой, образует строму органа.
Секреторная деятельность фолликулярных клеток (тироцитов) очень сложна и сводится к следующему.
1.Из аминокислот и солей, приносимых с кровью и проникаю щих в тироцит,-при активном участии рибосом, эндоплазматической сети, комплекса Гольджи образуется неиодированный тиреоглобулин, одной из аминокислот которого является тирозин. В виде мелких секреторных пузырьков он накапливается в апикальной зоне тироцитов и с помощью экзоцитоза поступает в полость фол ликула.
2.В полости фолликула в тирозин тиреоглобулина включаются последовательно атомы йода, которые образуются при окислении йодида, поглощенного из крови фолликулярными клетками. В ходе этого процесса последовательно синтезируются монойодтирозин, дийодтирозин, тетрайодтирозин (тироксин), трийодтиронин и на капливаются в коллоиде.
3.Тироциты своей апикальной поверхностью путем эндоцитоза поглощают (фагоцитируют) участки интрафолликулярного кол лоида, которые внутри цитоплазмы превращаются во внутрикле точные капли коллоида. Лизосомы соединяются с ними, после их расщепления образуются тиреоидные гормоны. Через базальную часть тироцита и базальную мембрану они поступают в общий кровоток, или в лимфатические сосуды (рис. 227, 228).
Таким образом, в состав гормонов, продуцируемых тироцитами, обязательно входит йод, поэтому для нормальной функции щито видной железы необходим постоянный его приток с кровью к щи-
310 |
311 |
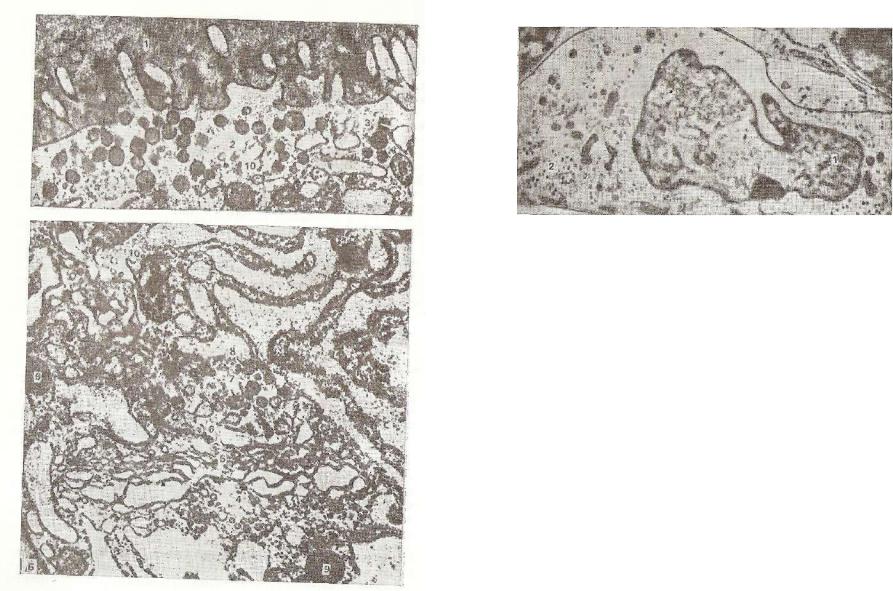
Рис. 227. Фолликулярная клетка щитовидной жвле'чм Гч™»,™™»™ |
||
рофотография): |
железы (электронная мик- |
|
А — апикальная часть клетки, обращенная к ТТПЪРТКППМЧ*. •, |
||
2 - апикальные гранулы; В — о р г а н е ™ т з д р т а ™ Ж ^ , : |
* Т » М и к р о в о р с и н к и : - |
|
лина; з — растянутые |
цистепны r K m S S ? *»™щ и е в |
с е к Ре Дии тиреоглобу- |
комплекс Гольджи; 5- |
тоансшотга!н£*яр*°й эндоплазматической сети; 4 L |
|
7 - секреторные гранулы? * - Жмл^нные nV3 bi^K»?°Se к р е т °Рн ы е гранулы;; |
|
митохондрии. |
окаймленные пузырьки; 9 g лазосомы; к? — |
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Околощитовидные железы развиваются из утолщения эндодер ма льного зачатка передних стенок 3-го и 4-го жаберных карманов; из мезенхимы образуются соединительнотканная капсула и про слойки желез.
Околощитовидные железы в количестве двух — наружного и и внутреннего эпителиальных телец — расположены около щито видной железы, а иногда и в ее паренхиме. Месторасположение этих органов у разных сельскохозяйственных животных сильно варьирует. Например, у крупного рогатого скота наружное тельце находится вблизи общей сонной артерии краниально от щитовид ной железы, внутреннее тельце — около дорсального края меди альной поверхности щитовидной железы; у лошади краниальное тельце лежит между пищеводом и краниальной половиной щито видной железы, каудальное тельце — на трахее. Паренхима около щитовидных желез построена из эпителиальных клеток — паратироцитов. Паратироциты образуют сложную систему переплетаю щихся тяжей, в которых различают их два основных типа: главные и оксифильные (ацидофильные) клетки. Имеются и как бы промежуточные формы. Между тяжами расположены тонкие про-
313
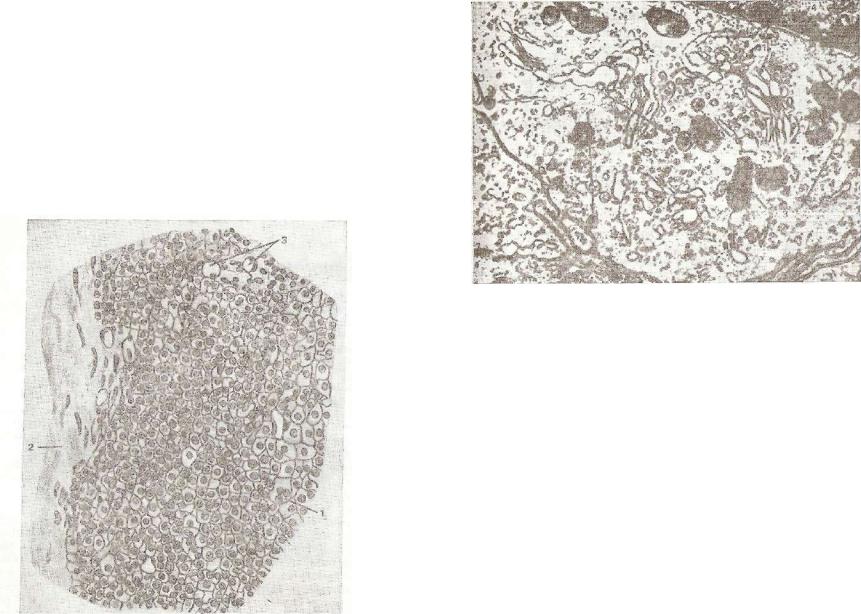
слойки рыхлой соединительной ткани с капиллярами и нервными волокнами (рис. 229).
Главные паратироциты составляют основную массу клеток. Они мелкие по размеру, полигональной формы, плохо окрашивают ся. Среди них удается различить более интенсивно окрашенные — темные клетки (плотные главные клетки) и менее окрашенные — светлые клетки (блестящие главные клетки). Главные паратиро циты содержат светлое ядро, незначительное количество зернис тости, воспринимающей специальные красители; гранулярную эндоплазматическую сеть, митохондрии, хорошо развитый комп лекс Гольджи. Секреторные гранулы покрыты мембраной и содер жат электроноплотную сердцевину (рис. 230).
Ацидофильные паратироциты крупнее главных. Их цитоплаз ма окрашивается кислыми красителями, содержит множество ми тохондрий и плотные ядра.
Рис. 229. Околощитовидная железа:
1 — паратироциты; 2 — соединительнотканная капсула: з — кроу веносные сосуды.
Главные паратироциты вырабатывают паратиреоидный гормон •(паратгормон). Он повышает содержание кальция и снижает со держание фосфора в крови; регулирует рост и регенерацию кост ной ткани; оказывает воздействие на проницаемость клеточных мембран и синтез АТФ.
Функция околощитовидной железы не зависит от гипофиза. Оксифильные и промежуточные паратироциты принято считать разновидностями главных клеток. Первые характеризуются высо ким метаболизмом, о чем свидетельствует обилие митохондрий.
Между тяжами паратироцитов может накапливаться вещество, сходное с коллоидом. Оно и окружающие его клетки формируют фолликулоподобные структуры.
Снаружи околощитовидные железы покрыты соединительно тканной капсулой. Она содержит тонкие нервные сплетения, от которых в паренхиму железы направляются нервные волокна. Васкуляризация околощитовидных желез обильна.
(НАДПОЧЕЧНИКИ
Надпочечники — это парные железы, каждая из которых построена из интерреналового и супрареналового органов, объединенных у позвоночных животных в единый орган. В составе надпочечника
314 |
315 |
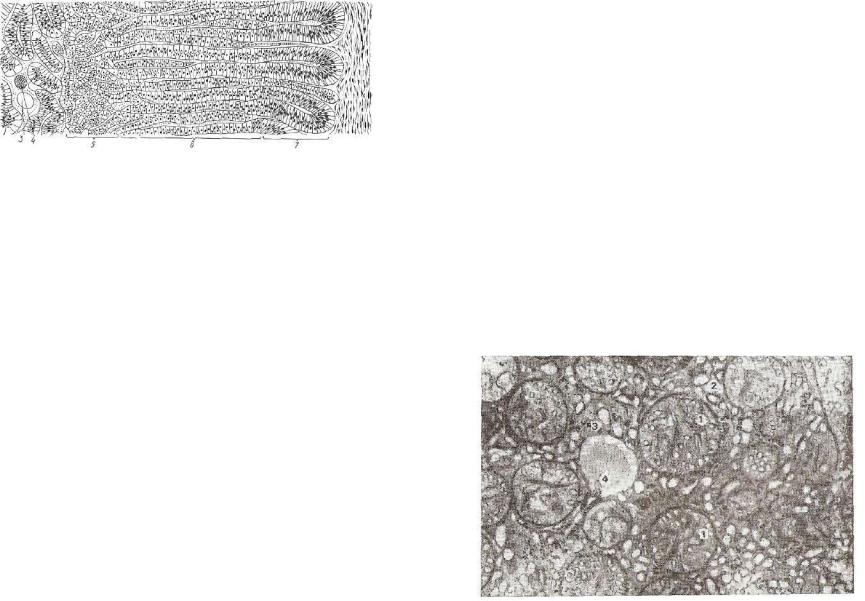
интерреналовый орган образует корковое, супр арена ловый орган — мозговое вещество (цв. табл. VIII).
Корковое вещество развивается из эпителиального утолщения целомической мезодермы, мозговое вещество, как и клетки симпа тических ганглиев, возникает из ткани нервных гребешков. Из мезенхимы образуется соединительная ткань органа.
Надпочечники имеют овальную или вытянутую форму и распо ложены вблизи почек. Снаружи они покрыты соединительноткан ной капсулой, от которой в глубь органа проходят тонкие прослой ки, направляющие вместе с кровеносными сосудами расположенно клеточных тяжей паренхимы.
Корковое вещество (кора) надпочечника лежит снаружи моз гового вещества, состоит из тяжей. В связи с их ориентацией, спе цифичностью строения и функции различают три зоны: клубочковую, пучковую и сетчатую (рис. 231).
К л у б о ч к о в а я з о н а построена из мелких секреторных кле ток, преимущественно цилиндрической (столбчатой формы). Они формируют скопления, в центре которых проходит соединительная ткань с кровеносными сосудами. Клетки содержат округлое ядро с более конденсированным хроматином и умеренно развитыми нит чатыми митохондриями, комплексом Гольджи, одиночными рибо сомами и полисомами. Большее развитие имеет агранулярная эндоплазматическая сеть в связи с синтезом гормонов стероидного типа.
В клубочковой зоне вырабатываются гормоны, регулирующие минеральный обмен, поэтому их называют минералокортикоидами. К последним относится альдостерон, регулирующий процесс реабсорбции натрия в почечных канальцах.
П у ч к о в а я з о н а представлена более крупными полигональ ными железистыми клетками, продуцирующими глюкокортикоиды — кортикостерон и кортизон. Клетки этой зоны образуют радиально ориентированные тяжи, между которыми проходят прослойки соединительной ткани с синусоидными капиллярами. В цито-
аодас ллсхил дириши уаавиты гладкая эндоплазматическая сеть, пузырчатые митохондрии с пластинчатой формой крист, лежащих вдоль или перпендикулярно продольной оси митохондрий (рис. 232). Клетки очень богаты липидами, поэтому на препаратах, получен ных классическими методами, цитоплазма имеет пенистый вид. Встречаются клетки с более темной цитоплазмой. Темные и свет лые клетки — это клетки, разные по своей функциональной актив ности.
Для с е т ч а т о й з о н ы характерны анастомозы и переплете ния ее клеточных тяжей. Клетки бедны липидами и более интен сивно окрашиваются. Хорошо развиты гладкая эндоплазматическая сеть, комплекс Гольджи; имеются рибосомы. Сетчатая зона выра батывает андроген. По химическому составу и характеру воздей ствия на организм он имеет сходство с мужским половым гор моном.
Отдельные скопления клеток коркового вещества образуют мел кие тельца, расположенные в грудной и брюшной полостях. Они носят названия интерреналовых телец и вместе с корковым веще ством образуют интерреналовую систему. Ее функция регулирует ся передней долей гипофиза. Исключение составляет клубочковая зона, действие которой не связано с гипофизом.
Мозговое вещество расположено в центральной части надпочеч ника. В его состав входят хромафинные клетки, симпатические нейроны, большое количество нервных волокон и синусоидных ка пилляров.
316
Хромафинные клетки — это видоизмененные нейроны симпати ческой нервной системы, которая регулирует их секреторную дея тельность. Они многоугольной формы, размер их до 30 мкм. Клет ки группируются в тяжи. Между ними проходят синусоидные ка пилляры, образующие обширные сплетения. Хромафинные клетки способны восстанавливать окислы хрома и других тяжелых метал лов, в цитоплазме у них откладывается буро-коричневый осадок, поэтому их и назвали хромафинными. В хромафинных клетках •слабо развиты гранулярная эндоплазматическая сеть, митохондрии. В полостях комплекса Гольджи накапливается электроноплотное вещество — предшественник секреторных пузырьков.
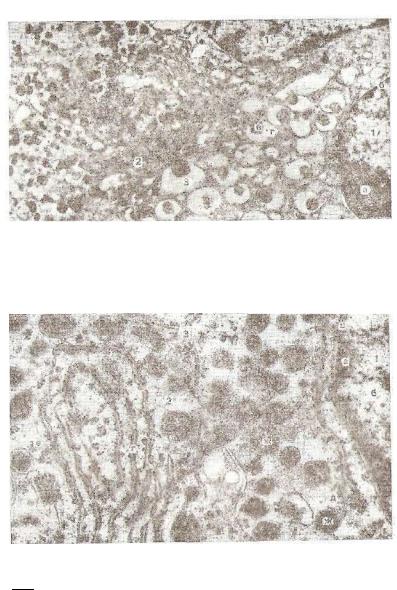
Различают две разновидности хромафинных клеток: норадреноциты и адренопиты. Первые вырабатывают норадреналин, вто рые адреналин. Содержавшиеся в норадреноцитах секреторные пузырьки, покрытые мембраной, характеризуются большим диа метром (100—300 нм), плотной сердцевиной (рис. 233).
Секреторные пузырьки адреноцитов отличаются меньшей элек тронной плотностью (рис. 234).
Норадреналин и адреналин относятся к катехоламинам. Нор адреналин является медиатором нервного возбуждения. Адрена лин — гормон, усиливающий работу сердца, регулирующий угле водный обмен. Для норадреналина эти свойства не присущи. Нор адреналин сокращает кровеносные сосуды и повышает давление, оказывает действия на нейросекреторную функцию гипоталамуса.
Мозговое вещество покрыто внутренней соединительнотканной капсулой, поэтому граница между корковым и мозговым веществом -отчетливо выражена.
Небольшие скопления клеток мозгового вещества надпочечника могут располагаться вблизи симпатических ганглиев, поэтому их называют параганглиями. Вместе с мозговым веществом они обра зуют хромафинную систему, функция которой не зависит от гипо физа и регулируется симпатической нервной системой. Этим обус ловливается наличие симпатических нейронов в составе мозгового вещества надпочечников.
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ ПТИЦ
Гипофиз птиц состоит из аденогипофиза и нейрогипофиза. В со ставе аденогипофиза отсутствует промежуточная доля. Аденогипофиз построен из дистальной и туберальной частей. В дисталь-
.ной части различают каудальную и нефальную (головную) доли.
Туберальная часть также неоднородна. В ней выделяют собст венно туберальную часть, портальную зону и внутреннюю туберальную часть.
Нейрогипофиз построен из нейральной доли, инфундибулярной -ножки и срединного возвышения.
Дистальная часть аденогипофиза представлена анастомозирующими между собой тяжами железистых клеток, окруженных кро веносными синусоидами. Среди тяжей могут встречаться кисты —
Рис. 233. Норадреноцит мозгового вещества надпочечника крупного рога того скота (электронная микрофотография):
1 —ядро- а —ядрышко; б —ядерная мембрана; г — цитоплазма; |
3 — секретов-, |
ные гранулы; в — плотная сердцевина; г — мембрана (снимок |
Плахотиной), |
Рис. 234. Адреноцит мозгового вещества надпочечника крупного рогатого-
скота (электронная микрофотография):
1 ядро; а — гетерохроматин; б — эохроматин; в — ядерная мембрана; 2 — цитоплазма; а — мембраны гранулярной цитоплазматической сети; д — рибосо мы; е — полисомы; ж — секреторные гранулы; 3 — плазмолемма (снимок ила— хотиной).
318
гштттттщтщтщтш*: я . : » . ^ ^ ^ н ^ »..^;.>..^ -•...;...,., 4 ....;.,,,,_ ,,,,..,. .. ч ... |
„. .,___.._ |
«•фолликулоподобные структуры. Их полости заполнены коллоидом. Число кист варьирует и в отдельных случаях может достигать -большого количества. Клеточные тяжи данной части гипофиза со стоят из хромофобных и хромофильных клеток. Последние харак теризуются различной способностью воспринимать красители, в связи с чем их классифицируют на базофильные, темные ацидо фильные, или Ai-клетки, и светлые ацидофильные клетки, или Аг-клетки. Базофильные клетки находятся преимущественно в |>астральном конце железы. В их цитоплазме содержится или голу бая, или синяя зернистость, или гомогенное вещество.
Зернистость ацидофильных клеток окрашивается кислыми кра сителями. Темные ацидофилы содержат крупную зернистость. Они 'чаще встречаются в каудальной доле, с возрастом их число умень шается. Имеются данные о том, что за б ч до овуляции их количе ство увеличивается, поэтому считают, что продуцируемые ими гормоны вызывают овуляцию.
В хромофобных клетках нет зернистости. Они обычно лежат в •центре клеточного тяжа и не контактируют с синусоидами. Как и у млекопитающих, эти клетки относят к малодифференцированным. Затем они могут превращаться или в базофильные, или в ацидо фильные клетки. Об этом свидетельствует наличие переходных форм.
Эпифиз у кур длиной около 2 мм и шириной 1 мм. Форма его треугольной пирамиды. Расположен он в небольшой полости между «большими полушариями и мозжечком. Снаружи эпифиз покрыт соединительнотканной капсулой, которая, проникая в глубь орга на, делит его на дольки. В междольковой соединительной ткани •часто встречаются скопления лимфоцитов.
У молодых животных эпифиз состоит из хорошо выраженных пузырьков, или фолликулов, заполненных коллоидоподобным ве ществом. В стенке фолликулов различают эпендимоциты, гипендимоциты, пинеоциты. Эпендимоциты являются наиболее многочис ленными клетками. По строению они очень сходны с эпендимными клетками центральной нервной системы. У них цилиндрическая форма; от их базального полюса отходит отросток, который закан чивается утолщением.
В период яйцекладки в этих клетках обнаружена суданофильеая зернистость. Между эпендимоцитами расположены гипендимоциты, а на периферии пузырьков находятся пинеоциты. Между пузырьками пространства заняты глиальными клетками.
У половозрелых животных фолликулы исчезают, падает гормонообразовательная активность клеток и они располагаются в виде клеточных скоплений.
Щитовидная железа у птиц состоит из двух топографически разобщенных желез. Они лежат в глубине краниального отдела грудной полости. Одна из желез находится под зобом на правой стороне пищевода, другая — слева на трахее.
Снаружи железы покрыты соединительнотканной капсулой, и, как у млекопитающих, они состоят из фолликулов и межфоллику-
320
лярнои ткани. Размеры фолликулов варьируют от 70 до 170 мкм» Внутри фолликулов расположен коллоид. Стенка фолликулов по строена из тиреоцитов, строение и гормональная функция которых обусловлены возрастными и сезонными изменениями. Во время линьки и в начале яйцекладки тиреоциты характеризуются наи большей активностью.
Электронно-микроскопическими исследованиями обнаружено, что тиреоциты покрыты плазмолеммой, которая на апикальной по верхности клетки образует многочисленные микроворсинки. Неко торые клетки снабжены одной центральной ресничкой, состоящей из девяти пар филаментов.
В тиреоцитах развиты комплекс Гольджи, эидоплазматическая сеть, содержатся гранулы различной электронной плотности. Обра зование плотных гранул связывают с функцией комплекса Гольд жи, а менее плотных—с эндоплазматической сетью. Предполагают, что эти гранулы являются предшественниками коллоида. Тиреоци ты лежат на базальной мембране.
Околощитовидная железа у птиц представлена двумя парами желез. Первая пара расположена справа у каудального конца щи товидной железы. Вторая пара находится от нее слева на расстоя нии 0,5—1,5 см. Железы овальной формы, образованы тяжами эпителиальных клеток, анастомозирующих друг с другом. Снаружи они покрыты соединительной тканью, обильно снабженной крове носными сосудами. Клетками эпителиальных тяжей являются: 1 — светлые клетки с водянистоподобной цитоплазмой, 2 — темные клетки с многогранным ядром, богатым хроматином; 3 — клеткипредшественники клеточным формам первого типа. Ядра их свет лые, овальной формы.
Оксифильных клеток у птиц не обнаружено. На строение и гормональную функцию клеток влияют возраст и сезонные изме нения. В период яйцекладки при интенсивном расходовании каль ция на построение скорлупы яйца количество светлых клеток значительно возрастает, превышая их в 10—20 раз.
Надпочечники у птиц также покрыты соединительнотканной капсулой. Они не разделены на корковое и мозговое вещество. Клетки образуют переплетающиеся тяжи и различаются по проис хождению. Часть тяжей развивается из целомического эпителия. Они называются главными, или кортикальными. Другие тяжи (промежуточные, или медуллярные) по развитию сходны с симпа тической нервной системой. Кортикальные тяжи более многочис ленны. Их клетки многогранной формы и расположены в два или три ряда обычно на периферии органа. Клетки промежуточных тяжей крупнее главных. Они содержат гранулы хромафинного ве щества, светлое ядро и базофильную цитоплазму. Цитоплазма кортикальных клеток богата митохондриями, вакуолями; слаборазвит комплекс Гольджи.
В главных тяжах различаются темные — неактивные и свет лые — функционально активные клетки.
Нервных клеток в паренхиме надпочечников нет. |
321 |
|
91 Заказ № 908 |
||
|
Г Л А В А 9
КОЖА И ЕЕ ПРОИЗВОДНЫЕ
Кожа — сложный и многофункциональный орган. Главная функ ция наружного покрова позвоночных — защита организма от вред ных воздействий окружающей среды. Сформировавшиеся в процес се эволюции модификации эпидермиса, который находится в прямом контакте с окружающей средой, существенны для поддержа ния постоянства внутренней среды в воде и в воздушной среде и защиты от опасностей окружения. Развитие кератинизированных придатков кожи, таких как. ороговевшие «зубы» у миноговых, че шуя, панцирь и коготки рептилий, чешуйки, коготки, перья и клюв птиц, чешуйки, когти, копыта, ногти, шерсть, волосы и рога мле копитающих, необходимо не только для защиты, по и для полового поведения, локомоции, хищничества, поддержания постоянной температуры тела и др. Пигментация, иммунные механизмы, ме- хано-, хемо- и терморецепторы также образуют существенные компоненты защитной системы и опосредуют приток информации из окружающей среды к телу. Защита обеспечивается также некератинизирующимися придатками кожи, такими как железы, отво дящими через выводные протоки на поверхность кожи ряд ве ществ. У птиц и млекопитающих такими веществами являются липиды, чтобы смазывать перья, шерсть и поверхность кожи, а у млекопитающих, кроме того, пот для регуляции температуры тела. Кожа участвует в обмене веществ, в процессах теплорегуляции организма, выделения, синтеза витаминов (витамин D) и др.
Хотя строение эпидермиса и его придатков различно у различ ных классов позвоночных, они обладают общими свойствами: 1) со стоят из эпителиальных клеток, происходящих из эктодермы, а под ними располагается дерма, происходящая из мезодермы; 2) содер жат популяцию герминативных клеток, дающих начало клеткам, дифференцирующимся непрерывно или циклами; 3) дерма влияет на все виды кератинизирующихся тканей, индуцируя и направляя ход дифференцировки.
В соответствии с видовыми особенностями животных кожа ха рактеризуется рядом специфических производных кожного покро ва: копыта травоядных животных, гребень птицы, рога, волосяной покров, молочные железы млекопитающих, перья у птиц и др.
КОЖА
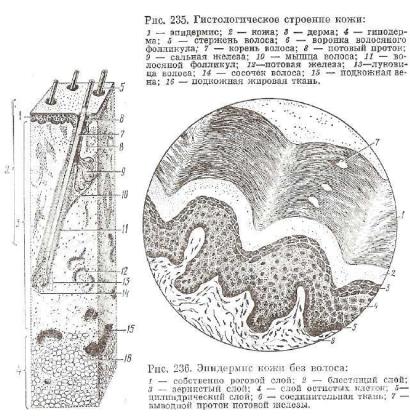
Кожа развивается из двух эмбриональных зачатков. Из эктодермы зародыша развивается наружный слой кожи—эпидермис (рис. 235). Глубокие слои кожного покрова — дерма и подкожная клетчатка — формируются мезенхимой, продуктом дифференцировки дерматомов зародыша.
Эпидермис кожи представлен многослойным плоским орогове^- вающим эпителием. Толщина его и степень ороговения специфич ны для вида животного, области тела и развития волосяного
322
покрова. Наиболее полно представлен эпидермис кожи в областях, не покрытых волосами. Процесс ороговения связан с накоплением клетками специфических фибриллярных белков — кератинов и вторичным их преобразованием. В эпидермисе можно выделить пять слоев: базальный, шиповатый, зернистый, блестящий и рого вой. Слои эпидермиса построены в основном из клеток, связанных с процессом ороговения, — кератоцитов (эпидермоцитов).
Б а з а л ь п ы й с л о й расположен непосредственно на базальной мембране. Состоит в основном из камбиальных эпидермоцитов и единичных меланоцитов (число меланоцитов относится к числу эпидермоцитов как 1:10). Граница эпидермиса с подлежащей сое динительной тканью дермы кожи неровная, что увеличивает пло щадь их контакта (рис. 236).
Эпидермоциты базального слоя цилиндрической формы. С клет ками своего слоя и расположенного выше шиповатого слоя они связаны десмосомами, а с подлежащей базальной мембраной полудесмосомами. Клетки этого слоя содержат многочисленные органеллы, в том числе и свободные рибосомы, о чем свидетельствует
21* |
323 |