

gistologia
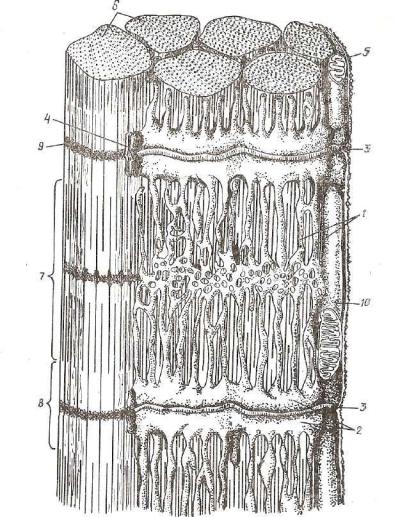
.pdfРис. 135. Гладкомышечные клетки (электронная микрофотография): ^Tij^5E2L* ~~ цитоплазма; 3 — митохондрии; 4 — комплекс Гольджи; 5 — эшюплазматическая сеть; 6 - толстые миофиламенты; 7 - плазмолемма (ув. 22 000) •
Б—8 — тонкие |
миофиламенты |
(ув. 90 000); В—9 — рибосомы: |
10 — гоаятаяттая |
|
вндоплазматическая сеть; |
л - |
агранулярная эндоплаКичеТкая сеть $ К Я о Т |
||
з ы Й к ! |
мембрана |
(ув. 46 000). Стрелками показаны |
пиноцитозные пу! |
|
Рис. 136. Гладкомышечные клетки (тангенциальный срез):
А—1 — ядро; г — цитоплазма; a — митохондрии; 4 — плазмолемма; 5 — толстые миофиламенты; б — коллаген; 7 — нервное волокно (ув. 14 000); В—8 — точки •сближения плазмолемм двух соседних миоцитов (ув. 20 000); В—9—внутренняя мем брана митохондрии; ю — наружная мембрана митохондрии; и — кристы; 12 — матрикс; 13 — миофиламенты (ув. 75 000). Электронная микрофотография.
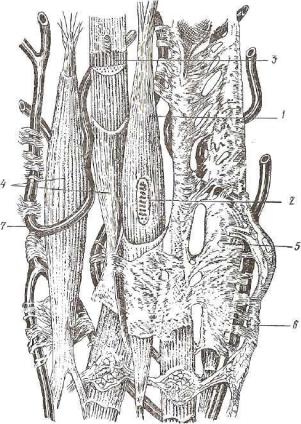
чительно выражены рибосомы, зернистая эндоплазматическая сеть (рис. 135). В цитоплазме миоцитов содержатся гранулы гли когена — энергетический резерв клетки.
Сократительным аппаратом миоцита являются миофиламен ты. Они расположены в цитоплазме на периферии клетки и име ют продольную ориентацию. Новыми цитологическими методами выявлены три их разновидности: миозиновые (толстые), актиновые (тонкие) и промежуточные. Диаметр миозиновых миофиламентов равен 17 нм, актиновых — 7 нм, промежуточных — 10 нм. Миофиламенты построены из сократительных белков. В световой микроскоп миофиламенты неразличимы.
186
В гладкомышечных клетках отсутствует поперечная исчерченность. Это объясняется ориентацией миофиламентов.
Плазмолемма миоцита, впячиваясь в цитоплазму, формирует многочисленные округлые образования, сходные с пиноцитозными пузырьками (рис. 136), примыкающие к саркоплазматической сети. Предполагают, что они участвуют в проведении нервных импульсов к сократительному аппарату миоцита, обусловливая этим сокращение. В основе последнего, очевидно, лежит, как и в исчерченной мышечной ткани, процесс скольжения актиновых и миозиновых (миофиламентов) нитей по отношению друг к другу.
Снаружи миоциты покрыты базальной мембраной. Из коллагеновых и эластических волокон вокруг каждого миоцита форми руется соединительнотканный футляр (эндомизий), который обеспечивает соединение миоцитов, не препятствуя процессу со кращения (рис. 137).
Если в мышечном пласте слабо развита соединительная ткань, то миоциты соединяются при помощи специальных межклеточ ных соединений типа десмосом (пятен слипания). Мышечный пласт построен так, что каждый его миоцит контактирует с не сколькими соседними клетками. Такое расположение способст вует более быстрому распространению нервного импульса, при этом в процесс сокращения вовлекается целая группа мышечных клеток.
Гладкомышечный пласт снабжен кровеносными, лимфатиче скими сосудами, нервными волокнами и окончаниями. В стенках многих внутренних полых органов гладкомышечные пласты фор мируют мышечные оболочки.
•ИСЧЕРЧЕННЫЕ (ПОПЕРЕЧНОПОЛОСАТЫЕ] МЫШЕЧНЫЕ ТКАНИ
Скелетная мышечная ткань — это сократительная ткань тулови ща, головы, конечностей, глотки, гортани, верхней, половины пи щевода, языка, жевательных мышц. Данную ткань относят к произвольной мускулатуре, так как ее сокращение контролиру ется волей животного.
Скелетная мышечная ткань развивается из миотомов сегмен тированного отдела мезодермы, а исчерченная мышечная ткань внутренних органов — из спланхнотома.
На ранней |
стадии развития миотомы состоят из плотно рас |
|||||
положенных |
мышечных |
клеток — м и о б л а с т о в . Это |
первая |
|||
стадия |
гистогенеза — м и о б л а с т и ч е с к а я . |
Цитоплазма |
мио |
|||
бластов |
имеет |
тонковолокнистое |
строение, |
свидетельствующее о |
||
развитии сократительных |
белков. |
Уже на этой стадии миобла- |
||||
сты способны к сокращению. Они интенсивно делятся и переме щаются клеточными потоками в участки расположения будущих мышц (рис. 138). Вскоре в цитоплазме миобластов можно разли чить единичные сократительные нити — миофибриллы, построен ные из сократительных белков. Ядра миобластов относительно крупные, овальные, с малым количеством гетерохроматина и хо-
187
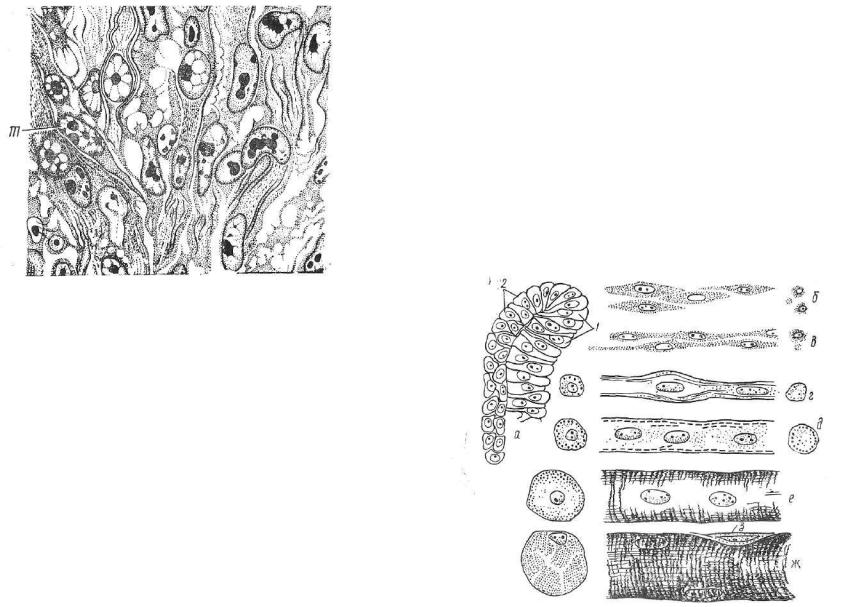
Рис. 138. Дифференцировка миобластов (т) в потоке клеток, выселяющихся из миотома.
рошо выраженными ядрышками. Они делятся интенсивнее кле ток, поэтому вскоре миобласты становятся многоядерными. Уве личиваясь в длину, они приобретают форму волокон — с и м п л а - стов .
В центре симпласта в ряд располагаются многочисленные ядра, на периферии интенсивно дифференцируются миофибриллы. Миосимпласты, очевидно, могут образовываться и путем слияния миобластов. Это вторая стадия гистогенеза. Она назы вается с т а д и е й м ы ш е ч н ы х т р у б о ч е к . Мышечные тру бочки, расщепляясь вдоль, формируют мышечные волокна. В по следних число миофибрилл резко увеличивается, многочисленные ядра перемещаются на периферию и располагаются под плазмолеммой. Волокно приобретает исчерченность. Это третья стадия
гистогенеза — с т а д и я м ы ш е ч н ы х |
в о л о к о н . К мышечным |
волокнам подрастает соединительная |
ткань с кровеносными со |
судами, нервами, дифференцируются нервные окончания. Соеди нительная ткань участвует в образовании наружной оболочки мышечного волокна и связывает между собой мышечные волокна (рис. 139).
Сведения о гистогенезе помогут понять строение скелетной мышечной ткани и разобраться в тех сложных изменениях, кото рые протекают в ней при физической нагрузке, тренировке, в условиях физиологической регенерации и патологии.
Процесс регенерации, протекающий в скелетной мышечной ткани, сходен с гистогенезом; в нем выявлены те же миобласти-
ческая стадия, стадия мышечных трубок и стадия мышечных: волокон.
Как следует из гистогенеза, у дифференцированной |
скелетной |
|||
мышечной |
ткани |
нет клеточного строения. Ее структурной и |
||
функциональной |
единицей |
является м ы ш е ч н о е |
в о л о к н о |
|
(рис. 140) |
в виде |
длинных |
цитоплазматических тяжей с закруг |
|
ленными концами, которые |
могут переходить в сухожилия. Дли |
|||
на волокон 10—100 мкм. Состоит мышечное волокно из сарко
плазмы (цитоплазмы) и |
многочисленных ядер, расположенных, |
на периферии. Само волокно покрыто сарколеммой (оболочкой).. |
|
Структурные компоненты |
саркоплазмы — сократительный аппа |
рат, органеллы, включения, гиалоплазма. Понять механизм сокра щения скелетной мышечной ткани можно лишь после знаком ства с тончайшей структурной организацией всех его компо нентов.
Сократительным аппаратом скелетного мышечного волокнаявляются продольно ориентированные м и о ф и б р и л л ы . По строенные из сократительных белков, они занимают большую: часть волокна, оттесняя ядра на периферию. Диаметр миофиб-
Рис. 139. Основные этапы эмбриогенеза скелетно-мышечной ткани:
а — клетки сомита (I — миотом, 2 — дермотом); б — миобласты; в — миосимпла сты; г — промиотуба; 9 — мышечная трубочка; е — незрелое мышечное волокно; ж — зрелое мышечное волокно; з — клетка соединительной ткани. Стадии б—ж показаны на продольном и поперечном разрезах.
189*
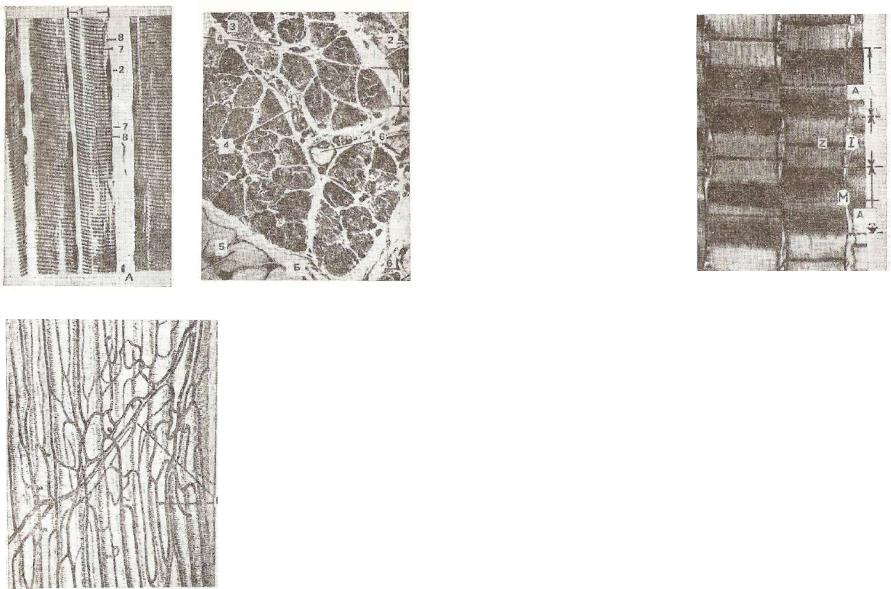
• —
Рис. 140. Исчерченная скелет ная мышечная ткань:
А — продольное сечение; Б — |
|
попереченое сечение; 1 — мышеч |
|
ное волокно; 2 — ядро мышечно |
|
го волокна; з |
— миофибриллы; |
4 — соединительная ткань пери- |
|
мизия; 5 — жировые клетки; б — |
|
кровеносный сосуд; 7 — анизо |
|
тропный диск; |
8 — изотропный |
диск; В — кровеносные сосуды мышечных волокон.
рил л около 1—2 мкм. Миофибриллы состоят из чередующихся
темных и светлых полос |
(дисков). Все |
светлые и |
все темные |
д и с к и м и о ф и б р и л л |
в одном мышечном волокне удержи |
||
ваются на одном уровне, в связи с чем |
волокно приобретает по |
||
перечную исчерченность. |
Продольная |
ориентация |
миофибрилл |
Рис. 141. Строение миофибрилл ис черченной скелетной мышечной тка ни:
А —• диск (анизотропный); J — диск (изотропный); Z-линия (телофратма); М-линия (мезофрагма) (по Хаксли). Элек тронная микрофотография.
может создавать продольную ис |
||||
черченность |
мышечного волок |
|||
на. |
|
|
|
|
В поляризованном свете тем |
||||
ные полосы |
(диски) |
обнаружи |
||
вают |
двойное |
лучепреломле |
||
ние — анизотропию, поэтому их |
||||
называют анизотропными, или |
||||
полосами А |
(дисками А). Свет |
|||
лые полосы изотропны, их назы |
||||
вают изотропными, или полоса |
||||
ми I |
(дисками I). |
В |
середине |
|
каждого |
I диска проходит |
темная зона — линия Z |
(телофраг- |
ма). В |
середине А диска |
проходит светлая зона — |
линия Н |
с темной линией посередине — линией М (мезофрагма) |
(рис. 141). |
||
Диски и линии были открыты очень давно с помощью оптического микроскопа. Они хорошо видны на изолированных миофибриллах, которые можно получить, расщепив мышечное волокно.
Структурной |
единицей миофибриллы является |
саркомер„ |
|||
В миофибрилле |
они расположены, следуя друг за другом. Сарко- |
||||
мер — это участок миофибриллы, состоящий из линии Z |
(для двух |
||||
соседних |
саркомеров), половины диска |
I, диска А |
с линией Нг |
||
половины |
следующего диска I, линии Z |
(для двух |
соседних сар |
||
комеров). Эти компоненты миофибрилл связывали с сокращения ми, однако их участие в этом процессе оставалось неясным. Элек тронно-микроскопические, гистохимические, биохимические иссле дования многое внесли в расшифровку функциональной морфо догии саркомера. Было установлено, что диск А состоит из более
толстых (диаметром 10 нм, длиной |
1,5 мкм) миофиламентов, |
диск I — из более тонких (диаметром |
5 им, длиной 1 мкм) мио- |
филаментов. Материалом для построения толстых миофиламентов служит белок миозин, а тонких — актин, тропомиозин В, тропин.
Актиновые и миозиновые миофиламеиты контактируют не ко нец в конец, а перемещаются по отношению друг к другу и в дис ке А образуют зону перекрытия. Участок А диска, состоящий только из миозиновых миофиламентов, называется Н линией и по сравнению с зоной перекрытия более светлый. Линия М — это место соединения толстых миозиновых миофиламентов в анизо тропном диске.
Линия Z состоит из Z-филаментов. В них выявлены белки' тропомиозин-В, а-актин. Z-филаменты формируют решетку, к ко-
190 |
191; |
|
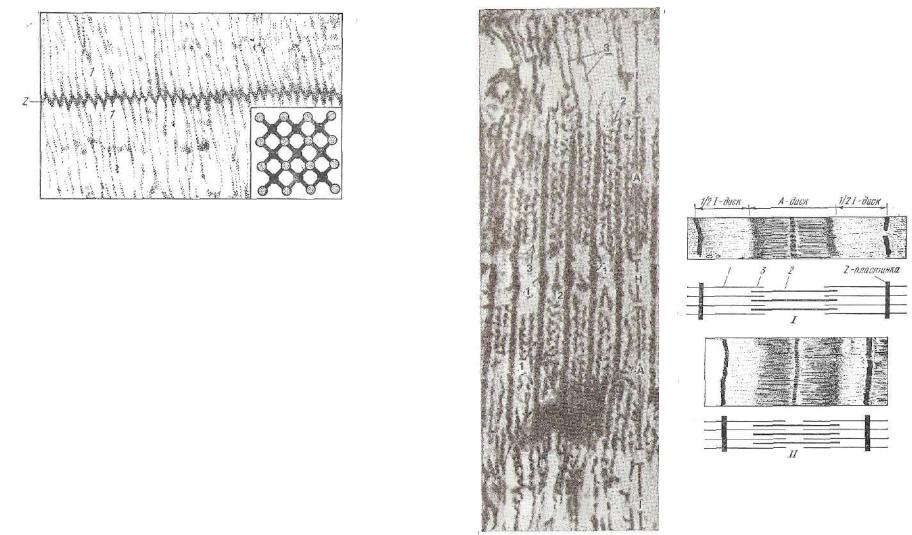
Рис. 142. Линия Z:
1 — прикрепление к ней тонких миофиламентов. Вставка вни зу поясняет прикрепление тонких миофиламентов к 2. Элек тронная микрофотография.
торой с обеих сторон прикреплены тонкие актиновые филаменты полосок I двух соседних саркомеров. Z линия проходит через всю толщину саркомера, а зона прикрепления тонких миофиламентов имеет зигзагообразный контур (рис. 142).
Таким образом, линии Z и М являются опорным аппаратом саркомера.
В строении сократительного аппарата при сокращении мышеч ного волокна наблюдаются следующие изменения: уменьшается длина саркомеров, так как тонкие (актиновые) миофиламенты полоски I при скольжении между толстыми (миозиновыми) филаментами полоски А сдвигаются к линии М диска А. Это приво дит к увеличению зоны перекрытия, формированию боковых мо стиков между актиновыми и миозиновыми миофиламентами (рис. 143), сокращению линий Н, сближению линий Z (рис. 144).
В гиалоплазме мышечного волокна хорошо развиты мито хондрии — органеллы клеточного дыхания. Они скапливаются между миофибриллами, вокруг многочисленных ядер, вблизи •сарколеммы, то есть в тех зонах, которые характеризуются зна чительной потребляемостью АТФ. Этим объясняется высокая метаболическая активность скелетного мышечного волокна.
Интенсивное развитие в мышечном волокне имеет незерни- •стая эндоплазматическая сеть (саркоплазматический ретикулум). Ее мембранные элементы расположены вдоль саркомеров и в виде терминальных цистерн окружают линии Z (рис. 145). Сар- •коплазматической сети присуща специфическая функция накоп ления иопов кальция, необходимых при сокращении и расслаб лении мышечного волокна.
Остальные органеллы (зернистая эндоплазматическая сеть, комплекс Гольджи и др.) развиты слабее и локализуются около ядер.
192
8 1 Рис. 143. Участок саркомера исчерчен ной мышечной ткани:
1 — толстые миофиламенты; 2 — попереч ные мостики; з — тонкие миофиламенты. •А — Vz Диска А; J — Vs диска I; H — зо на, состоящая только из толстых миофила ментов (по Хаксли).
Рис. 144. Саркомер поперечнополосатого мышечного волокна в расслабленном
(/) и сокращенном состоянии (II):
1 — тонкие нити; г — толстые нити; з — зона перекрытия.
Между миофибриллами находится значительное количество гранул гликогена (трофического) включения — материала для синтеза АТФ.
В цитоплазме мышечного волокна содержатся дыхательные ферменты, белок, миоглобулин — аналог гемоглобина эритроцитов; последний также способен связывать и отдавать кислород.
В мышечном волокне ядра расположены на периферии вбли зи сарколеммы. Они имеют овальную форму и варьируют по ко личеству от десяти до нескольких сотен. Гетерохроматин в виде крупных глыбок находится в относительно светлой нуклеоплазме. Ядра могут располагаться в виде цепочки, следуя друг за
13 Заказ № |
193 |
другом. Это результат амитотического деления — показатель ре активного состояния мышечного волокна.
Снаружи мышечное волокно покрыто оболочкой — сарколем мой, состоящей из внутреннего и наружного слоев. Внутренний слой — это плазмолемма, которая аналогична оболочке других тканевых клеток. Наружный — соединительнотканный слой со-
стоит из базальной мембраны и прилегающих к ней волокнистых структур. Плазмолемма образует систему узких канальцев, проei икающих внутрь мышечного волокна. Это система поперечных трубочек (Т-система). У млекопитающих системы Т-трубок распо ложены снаружи саркомеров на границе А и I дисков. У других классов животных она проникает в волокно на уровне линии Z. Зоны контакта системы поперечных трубок, саркоплазматической сети и терминальных цистерн называются т р и а д а м и . Они играют главную роль в продвижении волн деполяризации и аккумуляции ионов кальция. Триады видны только в электронный микроскоп.
Плазмолемма мышечного волокна, как и нервных волокон, электрически поляризована. В расслабленном мышечном волок не па ее внутренней стороне поддерживается отрицательный, на наружной стороне — положительный потенциал.
Примышечном сокращении волна деполяризации по нервно му волокну через нервное окончание перемещается на плазмолемму мышечного волокна, вызывая ее местную деполяризацию. Через систему Т-трубок, связанную с плазмолеммой, и триаду волна деполяризации оказывает влияние на проницаемость мем бран саркоплазматической сети, побуждая высвобождение в сар коплазму аккумулированных в ней ионов кальция. В присутст вии последних активируется расщепление АТФ, что необходимо для образования актомиозинового комплекса и скольжения акти ва овых миофиламентов по отношению миозиновых миофиламентов. Это вызывает укорочение каждого саркомера, а следовательно, миофибрилл и мышечных волокон в целом.
Важное место в этом процессе занимают молекулы толстых миофиламентов — м и о з и н а . Эти молекулы состоят из головки и длинного хвостика. Они при гидролизе АТФ, чему способствует АТФ-азная активность головок молекул миозина, вступают в связь с определенными участками молекул тонких миофиламен тов — а к т и н о м (см. рис. 143). Тонкие нити сдвигаются к цент ру саркомера, линии Z сближаются, увеличиваются зоны пере крытия, сокращаются линии Н анизотропных дисков миофибрилл ((см. рис. 144). Затем с участием АТФ актомиозиновые связи разрушаются, а миозиновые головки присоединяются к соседним участкам актиновых нитей, что способствует дальнейшему про движению миофиламентов по отношению друг к другу.
Если концентрация ионов кальция в саркоплазме снижается ш они перекачиваются в саркоплазматическую сеть, то сокраще ние мышечного волокна прекращается. Для этого процесса также необходима АТФ. Следовательно, и при сокращении, и при рас слаблении мышечного волокна расходуется АТФ, источником ко торой являются глюкоза, гликоген, жирные кислоты.
Сарколемма на концах скелетных мышечных волокон форми рует пальцеобразные выросты. Между ними находятся коллагеЁовые волокна соединительной ткани фасций и сухожилий, при крепляющие волокна к скелету.
13* |
195 |

Рис. 146. Развитие сердца:
А—В — поперечные разрезы вародышей |
|||
на трех |
послестадиях |
формирования |
|
трубчатой |
ванладки сердца; А |
— две |
|
парные закладки сердца; Б — их сбли |
|||
жение; В — их слияние в одну непар |
|||
ную закладку; 1 — эктодерма; 2 — эн |
|||
тодерма; 3 — париетальный листок ме |
|||
зодермы; 4 — висцеральный листок; 5— |
|||
хорда; 6 — нервная пластинка; |
7 — |
||
сомит; 8 — вторичная полость тела; 9 — |
|||
эндотелиальная закладка |
сердца |
(пар |
|
ная); Ю •— нервная трубка; 11 — по |
|||
лость сердца; is — эпикард; 13 — мио |
|||
кард; 14 — |
эндокард. |
|
|
Соединительнотканные во локна, расположенные снаружи базальной мембраны мышечного волокна, образуют эндомизий, который богат кровеносными со судами и нервами. Эндомизий соединяется с перимизием — оболочкой, покрывающей группу мышечных волокон. Перимизий нескольких мышечных пучков соединяется с эпимизием — са
мой наружной соединительнотканной оболочкой, объединяющей несколько таких пучков в мышцу — орган, характеризующийся специфическим строением и функцией.
Сердечная мышечная ткань. Этот вид мышечной ткани обра зует среднюю оболочку сердца, по характеру сокращения отно сится к непроизвольной, так как не контролируется волей живот ного. Развивается она из участка висцерального листка мезодер мы — миоэпикардиалыюй пластинки. Свое название эмбриональ ный зачаток получил в связи с тем, что из него развивается и другая оболочка сердца — эпикард (рис. 146).
Сердечная мышечная ткань |
состоит из мышечных клеток — |
к а р д и о м и о ц и т о в (сердечных |
миоцитов). Миоциты, соеди |
няясь друг с другом своими концами по длинной оси клеток, фор мируют структуру, сходную с мышечным волокном (рис. 147).
Границы |
между соседними |
миоцитами — это |
вставочные |
дис |
ки — аналоги линий Z, которые имеют прямые или ступенчатые |
||||
контуры. |
Вставочные диски |
обеспечивают механическую |
проч |
|
ность мышечного пласта и электрическую связь |
между кардио- |
|||
миоцитами. |
|
|
|
|
Различия в строении и функции миоцитов дали основание классифицировать сердечную мышечную ткань на две разновид ности: рабочую и проводящую. Первая составляет большую часть сердечной мышцы.
Кардиомиоциты на своей поверхности несут отростки илп анастомозы, так как с их помощью клетки соединяются друг с другом. Сердечные миоциты — это одноядерные и реже двуядер-
ные клетки. Их светлые ядра овальной формы расположены в центре клетки. Цитоплазма (саркоплазма) состоит из сократи тельных нитей — миофибрилл, органелл, включений и гиалоплазмы. Органеллы клеток локализуются у полюсов ядра. Хорошо развиты митохондрии, хуже комплекс Гольджи, саркоплазматическая сеть. Включения представлены многочисленными грану лами; гликогена и пигмента липофусцина. Количества последнего увеличиваются пропорционально возрасту.
Сократительный аппарат миоцитов так же, как в скелетной мышечной ткани, состоит из миофибрилл, которые занимают пе риферическую часть клетки. Их диаметр варьирует от 1 до 3 мкм. По своему строению миофибриллы сходны с таковыми скелетной мышечной ткани. Они также построены из анизотропных (поло сы А) и изотропных (полосы I) дисков. Этим обусловлена их поперечная иечерченность (рис. 148).
Элементы саркоплазматической сети окружают миофибрил лы. Характерное свойство сердечных миоцитов — отсутствие тер минальных цистерн, а поэтому и триад.
Плазмолемма кардиомиоцитов на уровень Z линий инвагинпрует в глубь цитоплазмы, образуя поперечные трубочки (Т-си- стему). От скелетной мышечной ткани они отличаются большим диаметром и наличием базальной мембраны, которая, как и сар колемму, покрывает их снаружи. Волны деполяризации, идущие с плазмолеммы, а также по Т-системе внутрь сердечных миоци тов, вызывают скольжение актиновых миофиламентов по отноше нию, миозиновых, обусловливая сокращение, как и в скелетной мышечной ткани.
196 |
197 |
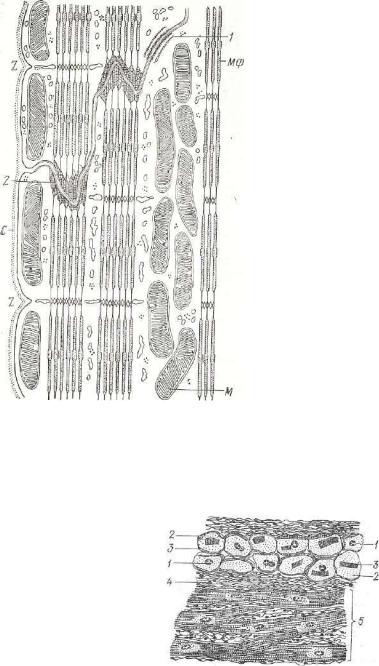
Рис. 148. Схема строения сердечной мышцы в об ласти ступенчатой вста вочной полоски:
С — сарколемма; М — ми тохондрии; МФ — миофиламенты; 1 — зона уплот нения на клеточной " обо лочке; г -• окончание мпофиламенты на плаамолемме; Z — полоска Z. Элек тронная микрофотография.
Проводящая мы шечная ткань также состоит из сердечных миоцитов, которые но сравнению с клетка ми рабочей мускула туры имеют больший диаметр, грушевид ную или удлиненную форму, богаты анасто мозами. Их светлые ядра с малым количе ством гетерохроматина и хорошо выра женным ядрышком локализуются в цент ре клетки. Цитоплаз ма богата гликогеном и бедна митохондри
ями, что свидетельствует об интенсивно протекающем в ней глико лизе и низком уровне окислительных процессов. Незначительно развиты рибосомы, саркоплазматическая сеть, система поперечных трубочек, мало миофибрилл. Последние занимают перифериче скую часть клетки и не имеют определенной ориентации, в связи с чем слабо выражена поперечная исчерченность. Так как миоциты содержат мало миоглобулина и внутриклеточных структур, то они окрашиваются слабее, чем клетки рабочей мускула туры (рис. 149).
Между собой кардиомиоциты проводящей мускулату-
Рпс. 149. Клетки проводящей мы шечной ткани сердца быка:
А — продольное, В — поперечное сечение; 1 — ядро; г — цитоплазма; S — миофибриллы; 4 — саркоплазма; 5 — рабочая мускулатура.
ры соединяются при помощи десмосом, а также щелевидных кон тактов, которые создают возможность прямого контакта ионов.
Эта разновидность сердечной мышечной ткани формирует систему, обеспечивающую проведение возбуждения.
СПЕЦИАЛИЗИРОВАННЫЕ СОКРАТИТЕЛЬНЫЕ ТКАНИ
Специализированные сократительные ткани построены из клеток и входят в состав потовых, слюнных, молочных желез. Сократительпые клетки этих желез в своей цитоплазме содержат миофиламенты, построенные из сократительных белков, и развива ются из эпителиальных клеток. Другие разновидности специали зированных сократительных тканей, имея нейроглиальное проис хождение и клеточное строение, суживают зрачок и располага ются в .радужной оболочке глаза.
НЕРВНАЯ ТКАНЬ
Нервная ткань — специализированная, высокодифференцировапная, формирует основную интегрирующую систему организма — нервную систему. Функция нервной системы определяется свой ством нервных клеток. Свое возбуждение они передают по цепи нейронов рефлекторных дуг на ткани рабочих органов, регулируя их взаимодействие в организме и с окружающей средой.
Нервная ткань состоит из нервных клеток — н е й р о н о в , вы полняющих специфическую для нервной ткани функцию нерв
ного возбуждения и проведения нервного импульса, и |
н е й р о- |
г л и и , обеспечивающей опорную, трофическую и |
защитную |
функции. |
|
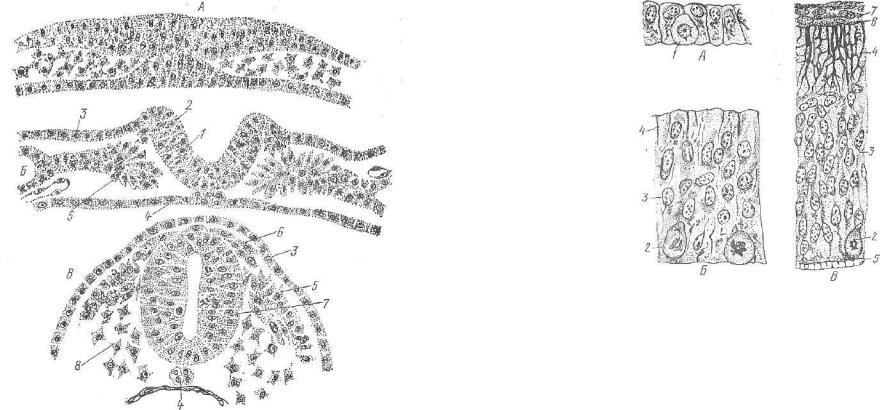
Развитие нервной ткани. Нервная ткань формируется из дор сального утолщения эктодермы — нервной пластинки, которая в процессе развития дифференцируется в нервную трубку, нейральные гребни (нервные валики) и нейральные плакоды (рис. 150).
В последующие периоды эмбрионального развития из нерв ной трубки образуется головной и спинной мозг. Нейральный гребень формирует чувствительные ганглии, ганглии симпатиче ской нервной системы, мелоноциты кожи, клетки системы APUD (энтерохромафиноциты, тироидные Са-клетки, хромаффинпые клетки мозгового вещества надпочечников, инсулоциты, выраба тывающие пептиды, обладающие активностью гормонов, и медиа торы).
Нейральные плакоды участвуют в формировании органов обо няния, слуха, чувствительных ганглиев (тройничного, лицевого, языкоглоточного и блуждающего нервов).
Нервная трубка на ранней стадии своего развития состоит из одного слоя призматических клеток. Последние, размножаясь, образуют три слоя: внутренний — эпендимный, средний — ман тийный и наружный слой — краевую вуаль (рис. 151). Первые
199
два слоя содержат активно размножающиеся клетки. Наружный слой образован отростками клеток двух первых слоев.
Впроцессе дальнейшего развития клетки внутреннего слоя продуцируют эпендимные клетки, выстилающие центральный ка нал спинного мозга. Клетки мантийного слоя дифференцируются на нейробласты, превращающиеся далее в нейроны, и спонгиабласты, дающие начало различным видам нейроглии (астроцитам, олигодендроглиоцитам).
Впроцессе дифференцировки нейробластов последовательно изменяется строение их ядер и цитоплазмы. В ядрах уменьша ется масса хроматина. В цитоплазме возрастает количество ри босом, формируются цистерны и канальца зернистой эндоплазматической сети, развивается комплекс Гольджи.
200
Рис. 151. Спинной мозг за родыша млекопитающих на разных стадиях развития:
А— нервная пластинка; В и
В— нервная трубка на более поздней стадии развития; 1 — митоз клетки нервной пластинки; 2 — митоз в эпендимном слое; з — ядерный (пла щевой) слой; 4 — наружный слой (краевая вуаль): 5 —• внутренняя пограничная мем брана; б — наружная погра ничная мембрана; 7 — мезен хима.
При световой микро скопии в цитоплазме на чинают выявляться фиб риллы (пучки нейрофиламентов и микротрубо чек). Нейробласты при нимают грушевидную
форму. Формируется отросток клетки - - нейрит. Позднее разви ваются дендриты, характерные для зрелых нейроцитов спинного мозга.
НЕРВНЫЕ КЛЕШИ
Нервные клетки (нейроциты, нейроны) различных отделов нерв ной системы характеризуются значительным разнообразием фор мы, размеров и функционального значения. В соответствии с функцией нервные клетки делятся на рецепторные (афферент ные, или чувствительные), ассоциативные и эффекторные (эффе рентные). Рецепторные нейроны под влиянием каких-либо воз действий внешней или внутренней среды организма генерируют нервный импульс и передают его па эфферентный нейроцит. По следний, возбуждаясь, передает его на ткань рабочего органа, по буждая последний к действию. Ассоциативные нейроциты обес печивают многообразные связи между рецепторными и эффекторными нейронами в составе рефлекторных дуг.
Нервные клетки различных отделов нервной системы весьма специфичны по размерам и форме. Например, диаметр перикариона клеток-зерен мозжечка равняется 4—6 мкм, а перикариона гигантских пирамидных клеток коры полушарий головного мозга достигает 130 мкм.
При большом разнообразии нервных клеток по форме общим морфологическим признаком нейроцитов является наличие от ростков, обеспечивающих их связь в составе рефлекторных дуг. В соответствии с локализацией отдельных нейронов длина их
20J
отростков весьма различна и колеблется в больших пределах—1 от нескольких микрон до 1—1,5 м.
Отростки нервных клеток по функциональному значению де
лят на два вида. |
Одни принимают нервное возбуждение и про |
||||
водят его к перикариону нейрона. |
В соответствии с тем что |
для |
|||
этих отростков |
большинства видов нейроцитов |
характерно |
вет |
||
вление, |
они получили название |
д е н д р и т о в . |
Другой вид |
от |
|
ростков |
нервных |
клеток проводит импульс от тела клетки и пе |
|||
редает его на другой нейроцит или на клетку рабочего органа — аксон (axis — ось), или нейрит. Все нервные клетки имеют толь ко один нейрит.
По количеству отростков нервные клетки |
делят па |
унипо |
||
л я р н ы е — с |
одним отростком, б и п о л я р н ы е — с |
двумя |
от |
|
ростками и |
м у л ь т и п о л я р н ы е — клетки, |
имеющие три |
и |
|
больше отростков (рис. 152).
Единственный отросток униполярных клеток всегда аксон, то
есть отросток, передающий |
нервный импульс на другую нервную |
|||
клетку. |
В организме |
млекопитающих животных униполярную |
||
форму |
имеют лишь нейробласты до периода образования дендри |
|||
тов. У |
биполярных |
клеток |
два отростка — нейрит и |
дендрит. |
У млекопитающих они характерны для органов чувств: |
сетчатки |
|||
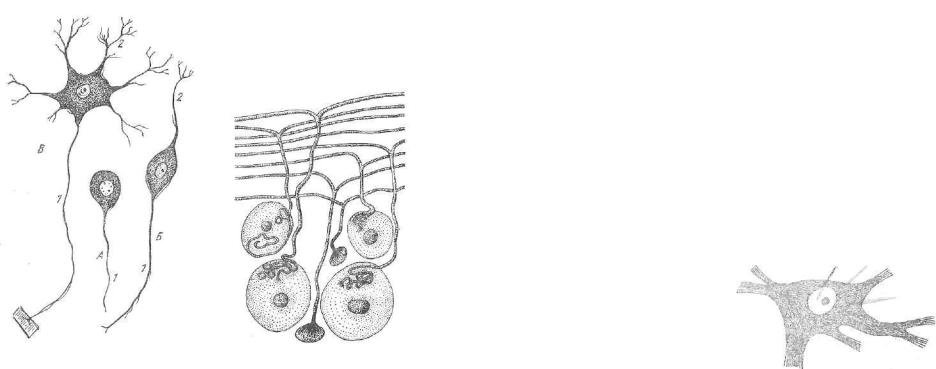
Рпс. 152. Нервные клетки: |
Рис. 153. Псевдоуниполярные |
А — униполярный нейрон; В — биполяр- |
клетки. |
рый нейрон; В — мультиполярный нейрон; |
|
I — нейрит; 3 —• дендрит. |
|
спинальных нервных узлов. Морфологически эти клетки характе ризуются тем, что от их перикариона отходит тяж цитоплазмы, имеющий форму отростка, который на некотором расстоянии от клетки Т-образно делится на дендрит, уходящий на периферию, и аксон, идущий в спинной мозг (рис. 153).
Ядра нервных клеток крупные, округлые или слегка овальные, располагаются в центре перикариона. Хроматин в ядре диспер гирован и образует небольшие скопления вблизи ядерной обо лочки. В ядрах мелких нейронов хроматина больше, что свиде тельствует 6 более низком уровне синтеза белков, необходимых для обеспечения целостности коротких отростков. Ядрышко крупное.
Цитоплазма нервных клеток характеризуется обилием раз личных органелл, что соответствует их высокой функциональ ной активности. При светооптическом анализе различными ме тодами исследования в цитоплазме нейронов выявляют нейрофибриллы, хроматофильное вещество, комплекс Гольджи, митохонд рии, центросому.
Поверхность |
клетки покрыта |
п л а з м о л е м м о й , |
которая |
|||
характеризуется |
возбудимостью и |
способностью проводить воз |
||||
буждение. |
Сущность этого процесса |
определяется закономерным |
||||
перемещением волны деполяризации |
(изменения разности потен |
|||||
циалов |
поверхностей плазмолеммы) |
|
по дендритам к перикариону |
|||
клетки |
и |
к нейриту. |
|
|
|
|
Ы е й р о ф и б р и л л ы — совокупность волокнистых |
структур |
|||||
цитоплазмы, элективно выявляющихся при обработке нервных клеток азотнокислым серебром. Электронно-микроскопическим анализом нейрофибриллярного аппарата установлено, что в его состав входят характерные для нейроцитов нейрофиламенты и
микротрубочки, формирующие в перикарионе |
нейроцитов густое |
|||
сплетение. |
В отростках |
нервных клеток они |
ориентированы па |
|
раллельно |
длине отростка, образуя пучки нейротубул |
и нейро- |
||
филаментов (рис. 154). |
|
|
|
|
Х р о м а т о ф и л ь н о е |
(базофильное) в е щ е с т в о |
выявляют |
||
в цитоплазме при окрашивании нервных клеток основными кра сителями (тионином, толуидиновым синим, крезиловым фиолето вым и др.) в виде базофильных глыбок различной формы и раз меров. Оно локализовано в периокариоие нейроцитов и в их дендритах, но отсутствует в аксональпых холмиках (область фор
мирования аксона) |
и в аксонах |
|
|||
(рис. |
155). |
Гистохимическим |
|
||
анализом установлено, |
что хро |
|
|||
матофильное |
вещество |
нервных |
|
||
клеток |
характеризуется |
высо |
|
||
ким содержанием РНК, а элект- |
|
||||
ронно-микроскопически |
показа |
|
|||
но, что глыбки хроматофильного |
|
||||
вещества соответствуют |
скопле- |
Р и с . 154. Нейрофибриллярный аппа- |
|||
нию параллельно |
расположен- |
рат нервной клетки (по Иванову). |
|||
203