

Ocherki_klinicheskoy_biokhimii
.pdf31
2.2. Метаболизм ДНК. Репликация
Проследим цепь событий. У нас есть клетка, которая живёт своей жизнью и которой, в общем – то, всё равно, что происходит вне её,она выполняет свою функцию, перекидываясь с товарищами цитокинами и метаболитами и меряется с ними размерами своих цитоподий. Внезапно, она получает «пинок»от старшего товарища по ткани в виде паракринных факторов роста (EGFR - epidermal growth factor receptor, IGF – 1 (инсулино – подобный фактор роста и проч.). Или поймала своим рецептором гормон, неважно. Ей приказано делиться.
Химический сигнал (словно восклицание соседки «а когда замуж), выводит из себя клетку, когда воздействует на её рецепторы к факторам роста. Серией реакций внутри клетки (обычно это каскад митоген – активных протеин – киназ, далее
MAPK – сигнальный путь), привёл к тому, что гистоны (после ацетилирования)
всеми фракциями отсоединились от ДНК, сделав её доступной для изнасилования в виде репликации, а клеточный цикл неожиданно, словно твоё отчисление из ВУЗа, достигло пресинтетической и синтетических фаз клеточного цикла. И мы имеем перед собой обнажённую ДНК в виде двойной спирали, из которой нужно сделать таких две. На первый взгляд миссия более чем невыполнимая, но, как и везде, нужно подходить к этому процессу последовательно. И с прелюдией.
Предлагаю не тянуть с этим.
В главных ролях: ДНК – полимераза; ДНК – хеликаза; ДНК – топоизомеразы; ДНК
– праймаза; ДНК – гираза; ДНК – связывающие белки.
Во второстепенных ролях: факторы, инициирующие транскрипцию ДНК, коих великое множество. Белки клеточного цикла, о которых мы поговорим позднее.
Этапы репликации ДНК. 1 этапинициация
2 этап – элонгация
3 этап -терминация
32
Репликация ДНК как таковая начинается с инициации.
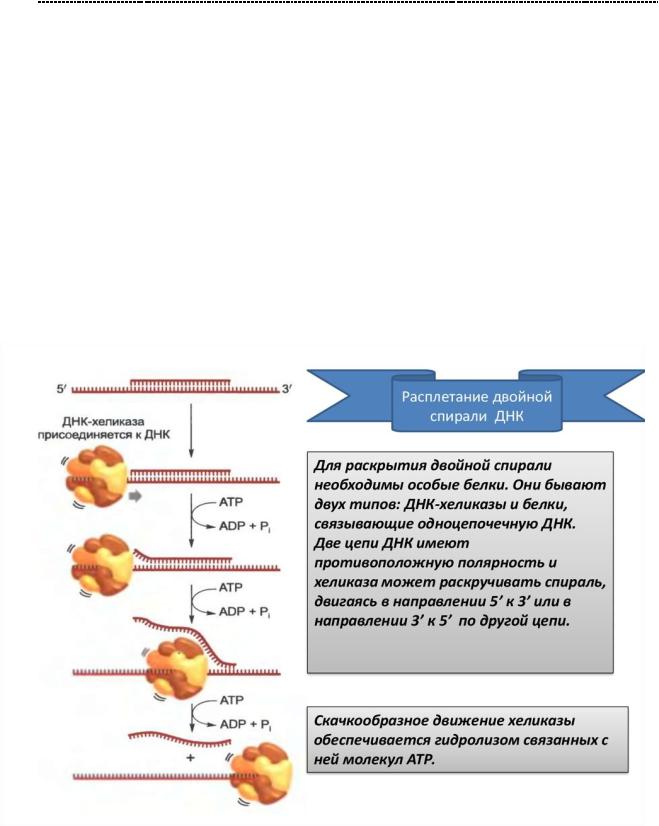
Уже известный нам товарищ, ДНК – хеликаза, присоединяется кточке инициации. (точке, характеризующейся множеством ТА - пар в структуре), к сайтусвязывания репликативного комплекса, после предварительной денатурации данного участка ДНК. Присоединившись на сайт связывания, Хеликаза движется от 5` к 3` - концу,
раскручивая ДНК по мере продвижения. Параллельно действуют ДНК – связывающие белки, удерживающие разъединяющиеся цепи ДНК от необдуманныхпоступков, иужеизвестныетопоизомеразы(ДНК–гиразаилиДНК
– топоизомераза 2), которая снимает топологическое напряжение (стремление к спирализации и замыканию в себе) с цепей ДНК.
(Первоисточник изображения неизвестен)
ДНК – полимераза не спешит работать. Она до последнего будет тянуть с синтезом нуклеиновой кислоты. Связано ли это с её тенденцией к
33
«созидательному созерцанию в виде прокрастинации или с её природным высокомерием, но она возьмётся за работутолько после того, как будет установлен
праймер. Праймер – это своеобразная РНК – затравка, это фрагмент РНК,
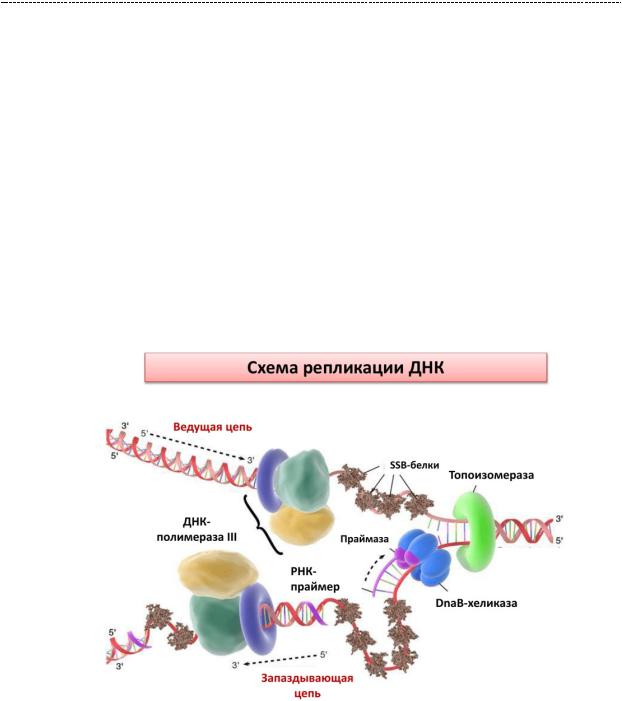
синтезированный РНК - праймазами. Он нужен для того, чтобы ДНК – полимеразе было что удлинять, ибо главная причина её долгого включения в работу – неспособность синтезировать РНК – фрагмент de novo, т.е. с нуля. Она может лишь удлинять имеющийся фрагмент. Сформированием праймеровсвязанпереход фазы инициации в фазу элонгации (удлинения) цепи. Фаза элонгации. ДНК – хеликаза связывается с РНК – праймазой, и это взаимодействие служит для синтеза лидирующей цепи ДНК взаимодействие.
(источник изображения неизвестен)
Синтез лидирующей цепи осуществляет ДНК – полимераза 3. Синтез на данной цепи осуществляется непрерывно, путём тупого присоединения дезоксирибонуклеотидов к 3 – штрих – концу предыдущего нуклеотида. Это называется от 5` к 3` - концу.
34
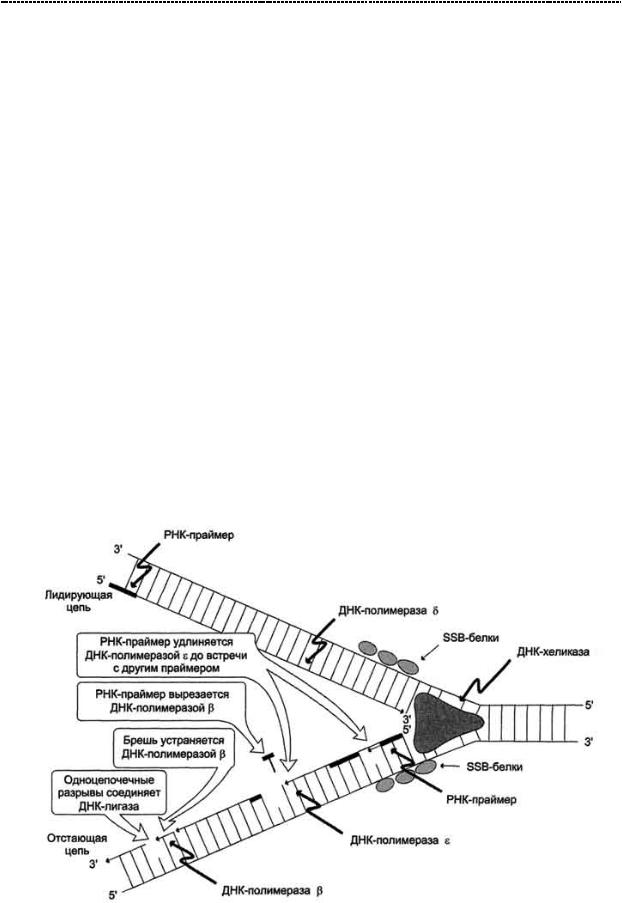
Элонгация. Но события разворачиваются так, что ДНК – полимераза 3
работает на «2 семьи» и синтезирует также и отстающую цепь, т.е. фрагментами Оказаки, по маленьким праймерам. Но что самое шокирующее:синтез происходит одновременно и скоординировано по времени. (а ты жалуешься, что тебе трудно совмещать учёбу с работой,). Секрет ДНК – полимеразы 3 прост: на отстающей цепи также работает комплекс ДНК – хеликазы + ДНК – праймазы (всё вместе это можноназватьпраймосомой)ДалеефрагментыОказакисшиваютсяДНК–лигазой.
Чтоб ты понимал, зачем нужны эти фрагменты Оказаки: ДНК – полимераза способна работать только в направлении от 5 штрих – до 3 – штрих конца цепи.
Отстающая цепь характеризуется противоположным направлением нуклеотидов и если бы её синтез тоже был непрерывным, репликативная вилка бы полностью распалась. А РНК – праймаза может синтезировать цепи в направлении 3`-5`. Она делает небольшой фрагмент, который затем просто удлиняется ДНК – полимеразой. А РНК – участок потом вырезается эндонуклеазой и РНК - азой.
Образовавшийся пробел достраивается ДНК – полимеразой (не 3, а уже другой) с
конца противоположного фрагмента Оказаки, и потом всё это склеивается ДНК -
- лигазой.
(иллюстрация - https://mylektsii.ru/2-96993.html)
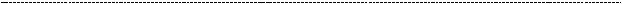
35
Получилось много текста, и резюмируя его, напомню основные моменты:
Для начала репликации нужно найти сайт связывания репликативного комплекса
ДНК – хеликаза необходима для разъединения цепей, иначе считать их невозможно
ДНК – праймаза и ДНК – полимераза необходимы для синтеза и удлинения нуклеиновой кислоты (ДНК), а ДНК – топоизомераза и ДНК – связывающие белки – для того, чтобы цепи ДНК не приняли «нечитаемую» конформацию
Одна дочерняя цепь синтезируется непрерывно, другая – прерывисто (с
помощью фрагментами Оказаки)
Нюансы и интересные факты:
- в ДНК есть также концевые фрагменты, состоящие из множестваповторяющихся пар нуклеотидов и не несущие существенной смысловой нагрузки. Эти фрагменты несут название теломеров. В ходе репликации эти концевые теломеры не воспроизводятся полностью. Преодолевая лимит Хейфлика (т.е. число делений без структурных потерь в цепях ДНК), теломеры укорачиваются до такой степени, что репликация в определённый момент становится невозможной. Но есть клетки, в
которыхестьтеломеразы–ферменты,достраивающиетеломеры.Такиетеломеразы есть в эмбриональных и опухолевых клетках.
- вбактериальныхклеткахпосле репликации хромосомыпереплетенымеждусобой в виде сыра «косичка», которое невозможно рассоединить простыми человеческими усилиями. Цепи вновь образованной ДНК переплетены нековалентно, но каждая из них замкнута ковалентными связами. Весь этот конгломерат цепей не распутать без участия ДНК – топоизомеразы IV (топоизомераза II типа). Они вносят кратковременный разрыв в обе цепи ДНК в одной хромосоме. Вторая хромосома может пройти через этот разрыв.
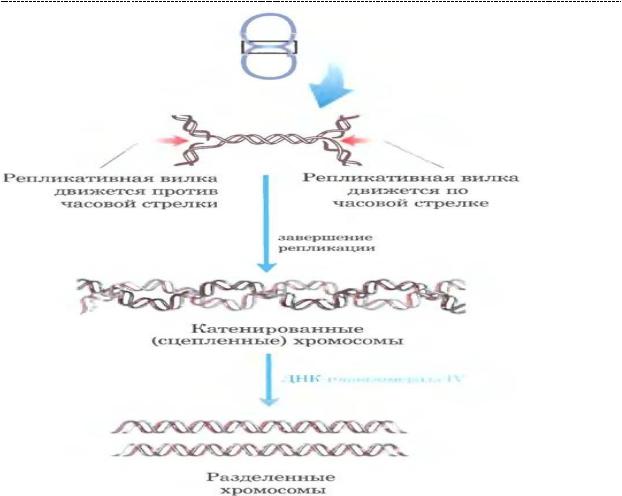
36
(источник иллюстрации – Основы биохимии Ленинджера, т.3, 2015 г)
- Топоизомераза IV является важной лекарственной мишенью для препаратов широкого (очень) спектра действия – хинолонов и фторхинолонов. Препараты избирательны по отношению к топоизомеразе IV и не проявляют интереса к топоизомеразам эукариот, что делает их применение очень удобным и подчас спасающим. Как ты понимаешь, если вновь образовавшиеся хромосомы похожи на сыр «косичку», адекватно считать с них информацию бактериальная клетка не может. И ей ничего не остаётся, как, в конечном итоге, погибнуть. Несмотря на кажущуюся селективность данных препаратов, их нельзя назвать абсолютно безопасными, но об этом ты узнаешь в своё время.
37
2.3.Синтез РНК и полипептидов.
2.3.1. Синтез РНК. Транскрипция. Синтез РНК в общих чертах напоминает
репликацию ДНК, с некоторыми нюансами:
-РНК–полимеразасамапосебесамодостаточна, иейненуженпраймердля начала работы.
-Субстратом для образования РНК являются рибонуклеотиды с аденином,
гуанином, цитозином и урацилом(вместо тимина).
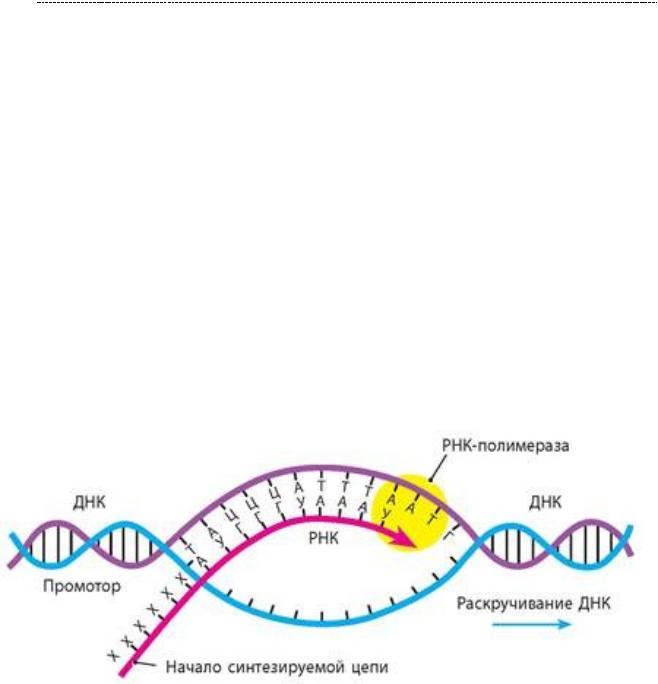
-Если при репликации копируется вся цепь ДНК, то в случае с транскрипцией читается лишь определённый участок ДНК, её ген, кодирующий определённый полипептид.
-Синтез РНК начинается при связывании РНК – полимеразного комплекса с промотором. Это участок ДНК, который сигнализирует о том, что «для синтеза данного полипептида нужно начать отсюда». Промотор богат ТАТА –
последовательностями нуклеотидных пар, за что получил кличку «ТАТА –бокс».
В остальном, даже стадии транскрипции повторяют за репликацией в виде инициации, элонгации и терминации. Но опять же, с некоторыми нюансами.
Инициация.Сначаланужнообразоватьтранскрипционныйкомплекс.Внего входят белки, обладающие хеликазной (т.е. «распутывающей нити») активностью;
белки, связывающие ТАТА – бокс; белки, стабилизирующие всю эту машину;
белки, связывающие РНК
– полимеразу 2 с ДНК в области её промотора (делающая это точно и безукоризненно), и, конечно же, РНК – полимераза 2 типа (да, у эукариот их 3) и
белки, которые её фосфолирируют.
Более того, есть даже белки в этой машине для синтеза РНК, которые обладают репаративной активностью, т.е. во время транскрипции осуществляют эксцизионную репарацию. В целом все эти белки можно обозвать факторами транскрипции, которым на Западе дали различные буквенные обозначения, но они нас сейчас малоинтересуют. Получается здоровая такая бюрократическая машина,
но с совсем не бюрократическими целями.
38
Инициация запускается в области промотора, но сама структура промотора не имеют ничего общего с кодирующей областью ДНК и с кодируемым ею полипептидом соответственно. Запускается она путём множественного фосфорилированияРНК–полимеразы2 участникамитранскрипционного тандема.
Элонгация. Удлинение цепи без регистрации и смс, путём простого присоединение нуклеозидтрифсофатов по направлению к 3 – штрих концу. РНК – полимераза 2 типа на стадии элонгации обладает нереально высокой активностью,
которая поддерживается связанными с нею белками (так называемые факторы элонгации.
Терминация. Когда весь здоровенный комплекс натыкается на сайты терминации,
РНК – полимераза дефосфорилируется и слезает с ДНК, выдавая готовую пре-
матричную РНК. Из неё уже можно получить что – то дельное.
(источник иллюстрации – интернет – портал https://history.vn.ua/pidruchniki/zadorozhnij-biology-9-class-2017-
rus/25.php)
Комментарии.
- У эукариот выделяют три РНК – полимеразы, но главную роль в генной экспрессии действительно играют именно РНК – Полимераза II. В свою очередь,
РНК – полимераза I образует пре – рибосомные РНК, которые участвуют в образовании рибосом – подпольных фабрик по синтезу белка. РНК – полимераза
39
III участвует в образовании специализированных РНК, функции которых до конца не охарактеризованы.
- РНК–полимеразытожеплачут. Нечасто используемый, ночрезвычайносильный антибиотик рифампиин (из группы ансамицинов) способен подавлять синтез бактериальных РНК путём связывания с В – субъединицей бактериальных РНК-
полимераз. Врезультатееёинактивации,непроисходитвысвобождениепромотора при транскрипции (по сути, не начинается транскрипция). Это позволяет использовать рифампицин в самых тяжёлых случаях – при стафилококковом остеомиелите, эмпиеме плевры и даже различных формах туберкулёза…
2.3.2. Процессинг как промежуточный этап на пути синтеза белка
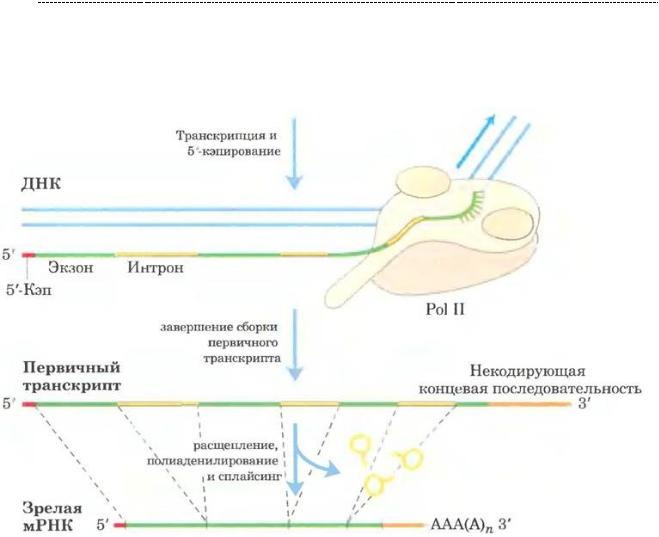
После образования первичного транскрипта (первичной, или пре – РНК)
происходит её обработка в виде процессинга. Он включает в себя:
-Кэпирование (присоединение 7 – метилгуанозина, связанного с 5` – концевым остатком мРНК). Функция этой метки – защита мРНК от рибонуклеаз, да и вцелом он имеет значение для инициации трансляции.
-Полиаденилирование – присоединение множества нуклеотидов с аденином на конец премРНК, имеет ту же цель, что и Кэпирование – защита от рибонуклеаз ядра и цитоплазмы (недолгая, но какая есть) и инициации трансляции. Данные участки способствуют правильной взаимоориентации малой субъединице рибосом и мРНК
-Сплайсинг. Это самая интересная часть, которая заключается в вырезании продуктов считывания интронов – не смысловых участков ДНК, не несущихв себе информации. Вырезание интронов происходит под действием сплайсосом (кстати,
РНК по своей природе). На выходе мы получаем пептид, состоящий из экзонов,но интересно даже не это. А то, что существует явление альтернативногосплайсинга.
40
Это тот вариант, когда могут нарезаться самые разные комбинации экзонов и получаться на выходе самые разные белки из одного первичного транскрипта, из одного гена. Это очень удобно для клетки.
(источник иллюстрации – Основы биохимии Ленинджера, т.3., 2015 г.)
2.3.3. Трансляция.
Процесс сложнее, чем кажется на первый взгляд. Как и на второй. В силу того,что для его осуществления клетке нужно много чего сделать.
В первую очередь, нужно перевести аминокислоты в такую форму, в которой они смогут участвовать в биосинтезе белка. Их нужно активировать. Для этого происходи присоединение к аминокислоте (любой) S-KoAгруппы с последующим присоединением к ней тРНК. Схематично и упрощённо, с отсутствием изображения стадии ацилирования аминокислоты, процесс выглядит следующим образом: