

2.Особенности репликации днк у про- и эукариотических организмов.
Репликация – это процесс самоудвоения молекулы ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплиментарности синтезируется дочерняя молекула ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре.
Полинуклеотидная цепь ДНК закручена в виде спирали и соединена с другой цепью, комплиментарной ей помощью водородных связей, образующихся между аденином и тимином (две связи), а также гуанином и цитизином (три связи). Нуклеотидв А и Т, Г и Ц называются комплиментарными. В результате у всякого организма число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых – числу цитидиловых. Благодаря этому свойству последовательность нуклеотидов в одной цепиопределяет их последовательность в другой.
Удвоение ДНК служит универсальным механизмом для воспроизведения генетической информации, необходимой для деления клеток.
Начинается репликация с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК.
Т.о. каждая нуклеотидная цепь выполняет роль матрицы для новой комплиментарной цепи. В результате получается две молекулы ДНК, у каждой из которых одна цепь родительская, а другая вновь синтезированная. Причем одна новая цепь синтезируется сплошной, а вторая – сначала в виде коротких фрагментов, которые потом сшивает ДНК-лигаза. В результате образуется две новые молекулы ДНК представляющие собой точную копию исходной молекулы.
Биологический смысл репликации состоит в точной передаче наследственной информации от материнской клетки к дочерним.
З.Типы энергетического метаболизма у бактерий.
По отношению к энергетическим источникам на две группы: фототрофы и хемотрофы.
Хемотрофные микроорганизмы используют для синтеза молекул АТФ энергию, освобождаемую при химических реакциях, фототрофные – световую энергию в процессе фотосинтеза. Образование молекул АТФ из АДФ может происходить двумя способами:
1) фосфорилирование в дыхательной или фотосинтетической ЭТЦ. Этот процесс у прокариот связан с мембранами или их производными, поэтому его называют М Ф-ем. При участии АТФ-синтазы:
АДФ + Фн → АТФ;
2) фосфорилирование на уровне субстрата. При этом фосфатная группа переносится на АДФ от вещества (субстрата), более богатого энергией, чем АТФ (субстратное фосфорилирование-СФ):
R ~ Ф + АДФ → R + АТФ.
В кл р-ции СФ не св. с М структурами и катализируются раствори-ми Ф промежуточного метаболизма.
У хемотрофов генерация Е в мол-ах АТФ сводится к 2 типам б/х реакций: окисления и восстановления. Окисляться м/о могут самые разнообразные орг. и неорг. ве-ва. Эти ве-ва - доноры электронов. Поскольку электроны не м. самостоятельно существовать, они обязательно д. б. перенесены на молекулы, способные их воспринимать, т.е. восстанавливаться. Эти молекулы - акцепторы электронов. Окисление субстратов происходит путём переноса электронов от донора к акцептору. При биологическом окислении чаще всего происходит одновременный перенос 2 эл-нов; при этом от субстрата отщепляются также 2 Н+ - это дегидрирование. У хемотрофов различают 3 типа Е м-зма: 1)аэробное дыхание или аэробное окисление; 2)анаэробное дыхание; 3)брожение.
Основным процессом энергетического метаболизма многих прокариот является аэробное дыхание, при кот. донором Н / эл-нов явл. орг. (реже неорг.) ве-ва, а конечным акцептором – О2. Осн. кол-во Е при аэробном дыхании образуется в ЭТЦ, т.е. в результате М Ф-я. Анаэробное – цепь анаэробных ОВР, кот. сводятся к окислению орг. / неорг. субстрата с исп. в качестве конечного акцептора эл-нов не О2, а др. неорг. ве-в (нитрата – NO3 -, нитрита – NO2- , сульфата – SO42-, сульфита – SO32-, CO2 и др.), а также орг. ве-в (фумарата и др.). АТФ образуется в ЭТЦ, т.е. в → р-ий М Ф-я, но в количестве <, чем при 1).
Брожение – совокупность анаэробных ОВР, при кот. орг. соед-ия служат как донорами, так и акцепторами эл-ов. К.пр., доноры и акцепторы эл-ов образуются из 1 и того же субстрата, подверг-ся брожению (например, из углевода). Сбраживанию м. подвергаться различные субстраты, но лучше др. - углеводы. АТФ при брожении синтезируется в → р-ий С Ф. Самое Е невыгодное – минимум Е.
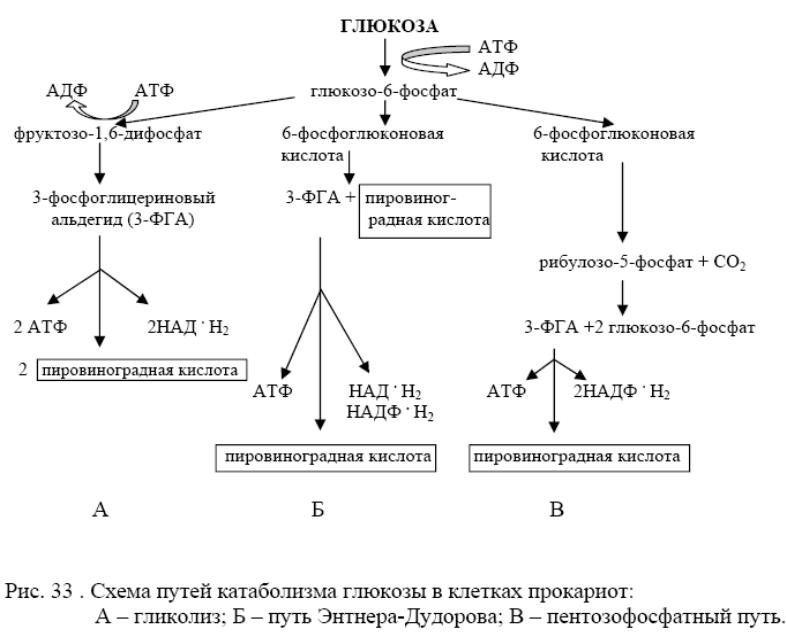
У б. 3 пути катаболизма глюкозы: 1) гликолиз; 2) окислительный ПФП; 3) путь Энтнера-Дудорова.
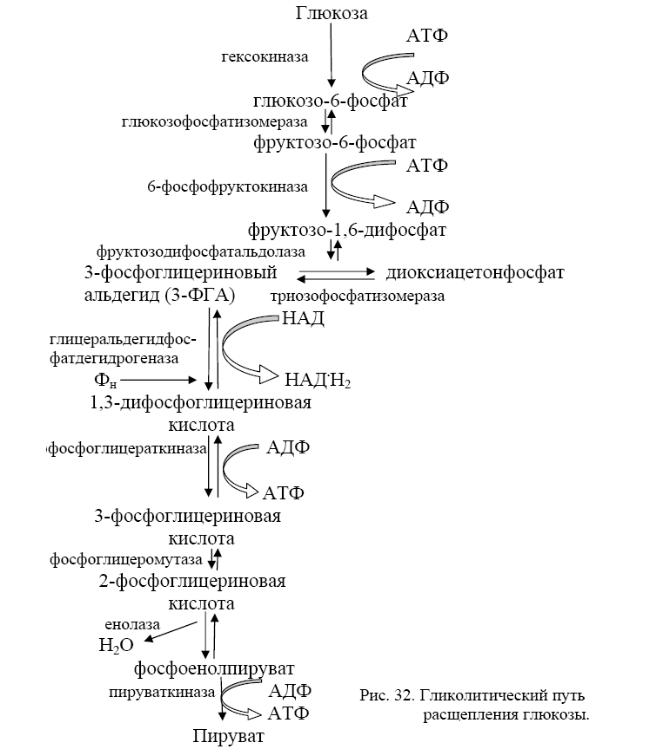
Все начинаются с того, что глюкоза в клетке сначала фосфорилируется при участии гексокиназы и АТФ как донора фосфата. Образуется глюкозо-6-фосфат, кот. представляет метаболически активную форму глюкозы в клетке и служит исходным пунктом для любого из 3 путей. Наиболее распространённый - гликолиз. При этом глюкозо-6-фосфат изомеризуется с помощью глюкозофосфатизомеразы и фосфорилируется далее в фруктозо-1,6-дифосфат, кот. затем расщепляется на 3-фосфоглицериновый альдегид (3-ФГА) и фосфодиоксиацетон (ФДОА). ФДОА под действием триозофосфатизомеразы → в 3-ФГА. Т.о., 1 мол. глюкозы → 2 молекулы 3-ФГА. На реакции превращения глюкозы в 3-ФГА затрачивается Е 2 АТФ. Далее - окисление каждой мол. 3-ФГА до 1,3-дифосфоглицериновой кислоты (1,3-ФГК). 1,3-ФГК – высокоэнергетическое соединение, содержащее макроэргическую фосфатную связь, – реагирует с АДФ (фермент фосфоглицераткиназа), отдавая эту группу, в результате чего синтезируется молекула АТФ. Т.о., Е, освободившаяся при окислении 3-ФГА, путём СФ оказывается аккум-ой в молекуле АТФ. Образуется 3-ФГК. Далее 3-ФГК под действием фосфоглицеромутазы в → 2-ФГК, из кот. в результате отщепления воды образуется фосфоенолпировиноградная кислота (ФЕП). Это также высокоэнергетический фосфат, с которого эта группа переносится пируваткиназой на АДФ, образуется молекула АТФ и ПВК. Это 2-ое фосфорилирование на уровне субстрата. Т.о., при распаде 1 молекулы глюкозы → 4 АТФ, в кот. аккум-ся освободившаяся при гликолизе Е. Поскольку в самом начале процесса на активирование глюкозы были израсходованы 2 АТФ, чистый выход АТФ на 1 молекулу глюкозы - 2 молекулы. С6Н12О6 + 2АДФ + 2Фн + 2НАД →2С3Н4О3 + 2АТФ + 2НАД*Н2.
Пентозофосфатный путь характерен для нек. Enterobacteriaceae, а также для гетеро-молочнокислых и нек. маслянокислых б. В этом цикле глюкозо-6-фосфат, образующийся путём активирования глюкозы молекулой АТФ, → ч/з ряд промежуточных р-ий в 6-фосфоглюконовую к-ту, кот. подвергается окислению и декарбоксилированию с образованием рибулозо-5-фосфата, СО2 и НАДФ*Н2. Рибулозо-5-фосфат включается в сложный цикл, → к обр. из его 2 молекул глюкозо-6-фосфата и 1 молекулы 3-ФГА. Глюкозо-6-фосфат может снова включаться в цикл. А 3-ФГА превращается гликолитическим путём в ПВК. С Е точки зрения этот путь в 2 раза < эффективен, чем гликолиз, т.к. на 1 молекулу глюкозы → только 1 молекула АТФ. Большое значение в том, что он обеспечивает клетки бактерий пентозами (рибулозо-5-фосфат), кот. - предшественники нуклеотидов и НК. + обр. 2 мол НАДФ*Н2, которые необходимы клетке для восстановительных реакций биосинтеза.
Путь Энтнера-Дудорова встречается у прокариот реже, чем др. Он характерен в основном для Ps. и уксуснокислых б. От ПФП он отличается тем, что 6-фосфоглюконовая к-та → в ПВК и 3-ФГА. 3-ФГА гликолитическим путём также → в ПВК. Из 1 молекулы глюкозы → 1 молекула АТФ, 1 молекула НАДФ*Н2 и 1 молекула НАД*Н2. Самый кратчайшим механизм расщепления С до ПВК. Т.о., важнейший продукт - ПВК, которая подвергается дальнейшим превращениям. Пируват занимает центральное положение в промежуточном метаболизме и м. сл. предшественником многих продуктов.
1) Аэробное дыхание. ПВК, образующаяся одним из 3 путей, окисляется до ацетил-КоА. Тут А пируватдегидрогеназы: CH3-CO-COOH + KoA~SH + HAД+→ CH3-CO~KoA + НАД*Н2 + CO2.
Ацетил-КоА - исходный субстрат ЦТК (ц. Кребса). В ц. Кребса включается 1 мол. ацетил-КоА, кот. + с оксалоацетатом (катализируется цитратсинтетазой). Продукты р-ии - лимонная к-та и свободный КоА. Цитрат с помощью аконитазы → в цис-акотиновую и изолимонную к-ты. Изолимонная к-та ч/з щавелевоянтарную к-ту → в α-кетоглутаровую к-ту, кот. подвергается дальнейшему декарбоксилированию. В итоге окисление ацетил-КоА в ЦТК даёт: 2 молекулы СО2, 1 молекулу АТФ и 8 атомов Н, из них 6 атомов Н на уровне пиридиннуклеотидов и 2 атома – на уровне флавопротеинов.
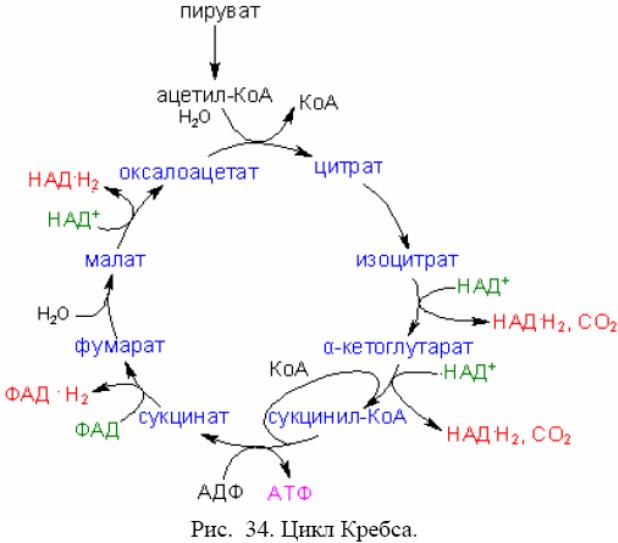
Т.о., ЦТК выполняет ф. конечного окисления орг. в-в. + клетку различными предшественниками (оксалоацетат, сукцинат, α-кетоглутарат и др.). У нек. б. он «разорван». Наиболее часто нету Ф этапа → α-кетоглутаровой к-ты в янтарную. В таком виде ЦТК не м. функционировать в системе энергодающих механизмов клетки. Осн. ф. «разорванного» ЦТК – биосинтетическая. НАД*Н2 и ФАД*Н2, образовавшиеся на разных этапах окисления орг. в-в, поступают в дыхательную цепь (ДЦ), кот. у б. находится в ЦМ, а у эукариот в М митохондрий. В ДЦ НАД*Н2 и ФАД*Н2 вновь окисляются до НАД и ФАД, а отщепившийся от них Н передаётся не <чем ч/з 5 переносчиков к концу цепи, где + с О2, образуя воду.
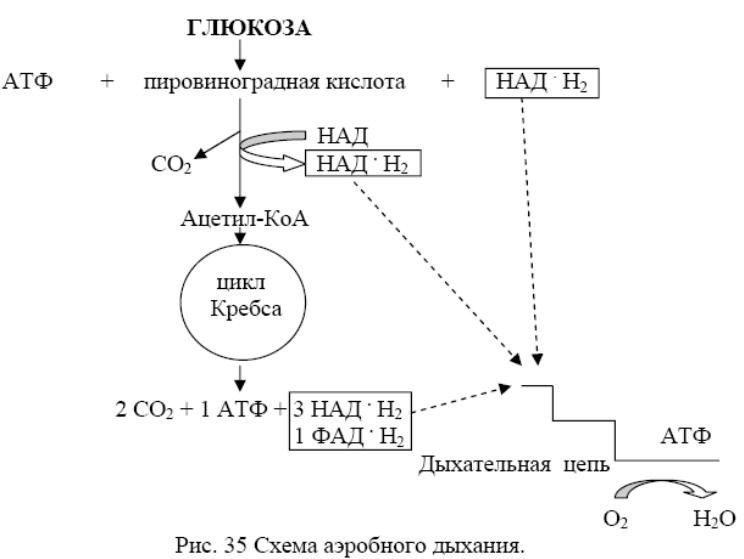
Переход Н по ДЦ состоит из ряда ОВР. В нек. из этих реакций выделяется достаточно Е для образования АТФ, и такой процесс носит название окислительного фосфорилирования (ОФ). В реакциях ОФ принимает участие специальная АТФ-синтаза, кот. катализирует → АДФ в АТФ. ДЦ м/о состоят из важнейших локализованных в М переносчиков атомов Н / эл-нов: флавопротеины, железосерные белки, хиноны и цитохромы.
Флавопротеины – коферменты, в состав которых входит витамин В2. В качестве простетических групп в них выступают флавинмононуклеотид (ФМН) или флавинадениндинуклеотид (ФАД). Флавопротеины осуществляют перенос атомов Н, т.е. являются дегидрогеназами. Дегидрогеназа, содержащая в качестве простетической группы ФМН, является НАДФ*Н2-дегидрогеназой. Это стартовый переносчик ДЦ, она осуществляет перенос Н с НАДФ*Н2 на последующие компоненты ДЦ. Дегидрогеназа, содержащая в качестве простетической группы ФАД, действует как сукцинатдегидрогеназа. Она катализирует окисление янтарной к-ты в фумаровую. Атомы Н от ФАД*Н2 поступают сразу на хинон.
Железосерные белки (FeS-белки) содержат железосероцентры, в кот. атомы железа св., с 1 стороны, с серой а.к. цистеина, а с другой – с неорг. сульфидной серой. Железосероцентры входят в состав некоторых флавопротеинов, например сукцинатдегидрогеназы и НАДФ.Н2 - дегидрогеназы, или же служат в качестве единственных простетических групп Б. ДЦ содержат большое число FeS-центров. Железосероцентры в зависимости от строения могут осуществлять одновременный перенос 1 или 2 электронов, что связано с изменением валентности атомов железа.
Хиноны – жирорастворимые соединения. У грам- б. хиноны представлены убихиноном (кофермент Q) или менахиноном. Хиноны липофильны и поэтому локализуются в липидной фазе М. Они переносят атомы Н. По сравнению с др. компонентами ДЦ они содержатся в 10 – 15-кратном избытке. Они служат «сборщиками» Н, поставляемого различными коферментами и простетическими группами в ДЦ, и передают его цитохромам. Т.о., они А в ДЦ на участке между флавопротеинами и цитохромами.
Цитохромы принимают участие на заключительном этапе цепи переноса эл-нов, Н они не транспортируют. К цитохромам эл-ны поступают от хинонов. В качестве простетической группы цитохромы содержат гем. Цитохромы окрашены; Различают цитохромы а, а3,b,c,o и др. Наиболее широко - цитохром с. Почти у всех, обладающих ДЦ. Конечные цитохромы ДЦ – это цитохромы а+а3 или цитохромоксидаза. Они передают эл-ны на О2, т.е. катализируют восстановление О2 до воды. В реакционном центре цитохромоксидазы содержатся помимо 2 гемов 2 атома меди. Т.о., ДЦ построена так, что одни компоненты ее переносят только атомы Н, а др. – только эл-ны. Флавопротеины и хиноны осуществляют перенос атомов Н, а FeS-белки и цитохромы – эл-нов. Переносчики атомов Н и переносчики эл-нов последовательно чередуются в ДЦ. По составу ДЦ различаются у разных м/о.
2) Анаэробное дыхание. При анаэробном дыхании конечным акцептором эл-нов в ЭТЦ – неорг. / орг. соединения, но не О2. Если конечными акцепторами эл-нов является SO42-, то процесс называют сульфатным дыханием, а б. – сульфатредуцирующими. Если - NO3- и NO2-, то нитратное дыхание / денитрификацией, а б. – денитрифицирующие. Если - СО2, то - карбонатным дыханием, а б. – метанобразующие. Если - фумарат, то - фумаратное дыхание. Б., способные к анаэробному дыханию, имеют укороченные ЭТЦ или ДЦ, т.е. они не содержат всех переносчиков. + в ДЦ анаэробов цитохромоксидаза заменена соответствующими редуктазами. У строгих анаэробов не функционирует ц. Кребса или же он разорван и выполняет только биосинтетические функции, но не Е. Основное количество молекул АТФ при анаэробном дыхании синтезируется в процессе М Ф-ия. По отношению к О2 б. – факультативные (денитрифицирующие б. и б., осуществляющие фумаратное дых.) / облигатные (сульфатвосстанавливающие и метанобразующие) анаэробами. Выход АТФ при анаэробном дыхании меньше, чем при аэробном дыхании, но больше, чем при брожении. Основные типы:
Нитратное дыхание - конечные акцепторы эл-нов - нитраты (NO3-) / нитриты (NO2-). В результате нитратного дыхания происходит восстановление NO3- / NO2- до газообразных продуктов (NО, N2О или N2). Способны только бактерии, у эукариот этот процесс не происходит.
Сульфатное - процесс окисления в анаэробных условиях субстрата (орг. соед. / Н2), при кот. конечный акцептор эл-нов - сульфат (SO42-). Происходит его восстановление до H2S. Б. - сульфатредуцирующие.
Карбонатное - процесс окисления Н2, при кот. конечный акцептор эл-нов - СО2. Б. - метанобразующие.
Фумаратное – отличается: роль конечного акцептора эл-нов в ДЦ – орг. в-во (фумаровая к-та); +этот способ запасания Е не является единственным для к.-л. гр б. Б., способные осуществлять это дых., являются хемоорганотрофами и им. способность к брожению. Это дополнительный механизм, позволяющим бактериям добывать повышенное количество энергии в анаэробных условиях.
3)Брожение – это способ получения Е, при кот. АТФ образуется в процессе анаэробного окисления орг. субстратов в реакциях СФ. При брожении продукты расщепления одного орг. субстрата м. одноt служить и донорами и акцепторами эл-нов. При сбраживании углеводов и ряда других в-в образуются (по отдельности или в смеси) этанол, молочная, муравьиная, янтарная к-ты, ацетон, СО2, Н2 и др. В зависимости от того какие продукты преобладают / являются особенно характерными различают спиртовое, молочнокислое (гетеро: лактат, спирт, СО2; гомоферментативное: тока лактат), муравьинокислое (муравьиная к-та, мало, но обр. только тут), маслянокислое (с. выгодное 3,3АТФ из 1 глю), пропионовокислое и другие типы брожения.
№23