

Изменения в природных биологических системах - Федоров В.Д
..pdfНесомненно, могут быть предложены и другие способы оценки поведения отдельных переменных системы.
Независимо от того, каким способом произведена оценка стабиль5 ности отдельных переменных природных экосистем, можно с боль5 шой долей вероятности ожидать, что при ранжировании найденных значений Si вся полученная совокупность оценок образует бимодаль5 ное распределение. Экспериментальное подтверждение бимодально5 го распределения оценок стабильности отдельных переменных, харак5 теризующих экосистему пелагического планктона, было получено нами ранее (Федоров, Соколова, 1972). Вследствие того, что  и
и  нор5 мированы
нор5 мированы  , найденные значения Si расположе5 ны в интервале отрезка единичной длины, образуя две группы оценок (бимодальное распределение значений). При этом одна из совокуп5 ности оценок, полученных одним из способов, относится к показате5 лям, которые характеризуют скорость процесса (интенсивность функ5 ции) и, следовательно, эти оценки могут быть выражены производ5 ной во времени. Назовем их функциональными, полагая, что
, найденные значения Si расположе5 ны в интервале отрезка единичной длины, образуя две группы оценок (бимодальное распределение значений). При этом одна из совокуп5 ности оценок, полученных одним из способов, относится к показате5 лям, которые характеризуют скорость процесса (интенсивность функ5 ции) и, следовательно, эти оценки могут быть выражены производ5 ной во времени. Назовем их функциональными, полагая, что
(4)
Вторая совокупность группирует оценки показателей, которые могут быть рассмотрены как некоторый итог процессов (функций) к моменту наблюдений и, следовательно, такие оценки могут быть выражены интегралом во времени. Назовем их структурными, полагая, что
(5)
Важнейшая функциональная характеристика экосистемы –продук5 тивность, отражающая скорость образования органического веще5 ства (биомассы) отдельными популяциями видов, образующими сообщество. Важнейшая структурная характеристика экосистем – биомасса популяций. Разница между величинами биомассы и про5 дуктивности аналогична различию оценок, получаемых при опреде5 лении концентрации хлорофилла (в мкг/м2) в текущий момент времени
искорости изменения его концентрации (мкг/м2 в сек, час, сутки). Возникает задача: как на основе многочисленных отдельных оце5
нок независимых переменных можно количественно оценить целос5 тные свойства системы? Или, сформулировав задачу более абстракт5 но, как перейти от многих частностей к целостной оценке общности?
При этом следует сразу признать, что оценки, получаемые по формулам (4) и (5), характеризуют среднюю стабильность отдель5 ных переменных и к устойчивости экосистемы непосредственного отношения могут не иметь. Действительно, легко представить себе, что устойчивость системы в целом обеспечивается благодаря неус5
171

тойчивости образующих частей. Могут быть выдвинуты и другие правдоподобные постулаты, пригодные для логического анализа и экспериментальной проверки. Во всяком случае, попытка Паттена (Patten, 1961) охарактеризовать стабильность экосистемы средним значением стабильностей отдельных переменных не представляется сколько5нибудь убедительной.
Поэтому отыскание решений для сформулированной задачи ока5 зывается делом нетривиальным, хотя и небезнадежным в принципе. На пути отыскания решения поставленной задачи разделение пока5 зателей биотической компоненты экосистемы на функциональные и структурные нам представляется весьма плодотворным, поскольку при этом возникает возможность сопоставления средних оценок (или положений модального класса) для двух совокупностей получен5 ных оценок – Sф и Sc.
Нетрудно заметить, что сопоставление оценок Sф и Sc приводит к введению некой меры, характеризующей внутреннее состояние системы, и эта мера, как будет показано ниже, определяет гомеос5 таз системы.
3. ГОМЕОСТАЗ БИОЛОГИЧЕСКИХ СИСТЕМ
Первоначально термин «гомеостаз» был введен в 1932 г. Кенне5 ном (Cannon, 1932) для обозначения механизмов поддержания рав5 новесия внутренней среды (например, постоянства количества сахара в крови, постоянной температуры тела у теплокровных животных), непрерывно нарушаемого изменяющимися условиями внешнего мира и непрерывно восстанавливаемого внутренними специальными ме5 ханизмами с обратной связью действия. Эти механизмы находятся под управлением вегетативной нервной системы и, следовательно, присущи всем живым системам с центральной подсистемой управ5 ления, т.е. системам а5типа, или просто а5системам.
Много позднее (Федоров, 1970а), анализируя особенности орга5 низации а5 и r5систем, мы обратили внимание на существование принципиального различия между ними. Так, для а5систем функци5 ональные перестройки направлены на поддержание, в конечном счете, постоянства структурных характеристик (например, сохра5 нение специфического внешнего облика систем за счет функциони5 рования многочисленных механизмов регуляции обмена веществ). У r5систем структурные перестройки направлены на поддержание постоянства функциональных характеристик. Так, например, изме5 нение соотношения свето5 и тенелюбивых форм, тепло5 и холодолю5 бивых видов позволяет поддерживать постоянство продукционных характеристик в течение вегетационного периода при варьировании температурных условий и освещенности. Таким образом, наличие экологических механизмов, периодически поддерживающих посто5
172

янство функциональных характеристик r5систем в пределах опре5 деленных границ, позволяет применить к ним концепцию гомеоста5 за после ее обобщения и формализации.
Будем считать, что понятие гомеостаза распространяется на ме5 ханизмы поддержания постоянства одних характеристик благодаря непостоянству других. Это означает, что возникновение относительно устойчивых свойств в одной части системы обеспечивается благода5 ря возникновению дополнительной неустойчивости других ее свойств.
Таким образом, гомеостаз – механизм регуляции, упорядочиваю5 щий во времени изменение свойств системы в направлении устойчиво5 сти групп характеристик, относящихся либо к процессам (r5систе5 мы), либо к их результатам (а5системы). В экологических системах, в частности, возникновение дополнительной зоны непостоянства связа5 но с изменением относительного обилия отдельных популяций, т.е. с изменением удельной доли биомасс видов при сохранении постоянства общей скорости ее образования организмами однотипного сообщества.
В предыдущей работе (Федоров, Соколова, 1972) мерой гомеос5 таза (G) живой компоненты экосистемы служило отношение
(6)
Однако по соображениям чисто практических удобств, связан5 ных с введением нормирования 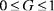 , при пользовании показа5 телем стабильности меру гомеостаза лучше рассчитывать из выраже5 ния (7), если значения Si определяются по формуле (3):
, при пользовании показа5 телем стабильности меру гомеостаза лучше рассчитывать из выраже5 ния (7), если значения Si определяются по формуле (3):
при |
(7) |
а при расчете оценок стабильности Si – по формулам (1) и (2):
при |
(8) |
Таким образом, в живой системе различным переменным может быть приписана различная абсолютная мера изменчивости, в зави5 симости от выбранного способа оценки Si.
Закономерно возникает вопрос: что является причиной изменчи5 вости отдельных характеристик? Таких причин две.
Во5первых, внутренние причины, обусловленные особенностями взаимодействия слагающих систему элементов, отражают правила соподчинения элементов и связей между ними. Таковы, например, колебания численностей в системе двух популяций при взаимоотно5 шениях между ними типа «хищник – жертва». Важнейшая особен5 ность вызываемых внутренними причинами изменений – их флюк5 туирующий характер, т.е. изменение их с переменной амплитудой и
173

частотой в пределах некоторой области, которая является нормой при обычных условиях существования системы. Таким образом, норма относится, прежде всего, к отдельным функциям как первич5 ным показателям, характеризующим особенности как биотической, так и абиотической компонент экосистемы. Несомненно, однако, что должна существовать и «норма отдельных целостных свойств систе5 мы» и даже всей системы в целом.
Отклик экосистем на внесистемные воздействия расширяет поня5 тие нормы, поскольку внесистемные факторы образуют изменяющий5 ся фон, на котором действуют внутренние причины. Поэтому «норма» либо должна быть приурочена к отдельным периодам, характеризу5 ющимся относительным постоянством фона (вероятно, можно гово5 рить о норме периода, например, о норме биологического лета), либо должна охватывать крайние пределы возможного изменения фона, что, естественно, расширяет норму всех регистрируемых показателей (таким образом, можно говорить о норме существования экосистемы).
Во5вторых, внешние причины, обусловленные особенностями воздействия возмущающих факторов окружающей среды, способ5 ны вызвать отклонения заданных переменными свойств системы за пределы нормы.
Таким образом, именно возмущающие причины ставят вопрос о допустимой мере отклонений от нормы, в пределах которой экосисте5 ма продолжает оставаться самой собой, т.е. сохраняет свое лицо, обус5 ловленное особенностями своей функциональной, пространственной, генетической и экологической структуры. Иными словами, система приобретает способность противостоять внешним воздействиям, бла5 годаря свойству ликвидировать их последствия в результате упорядо5 чивающих положение механизмов гомеостаза. Эта способность приво5 дит нас к понятию устойчивости как целостной характеристики сис5 тем, выраженной в способности противодействовать возмущениям.
Легко заметить, что это общее и, кажется, достаточно ясное тол5 кование устойчивости малоконструктивно, т.е. оно не способствует установлению принципов исследования устойчивости, не намечает конкретных путей подхода к измерению этой способности системы противостоять возмущениям. Поэтому обратимся еще раз к поня5 тию нормы и меры отклонений, чтобы придать понятию устойчиво5 сти «рабочий вид».
Только взгляд на состояние системы как на поддержание некото5 рой нормы свойств, обусловленных в свою очередь нормой отдель5 ных переменных, позволяет поставить вопрос об устойчивости в плос5 кость установления допустимой меры отклонений от нормы некото5 рых ее свойств при воздействии конкретных возмущений среды.
Тогда определим устойчивость систем как допустимую (без рис5 ка разрушения системы) меру отклонений заданных свойств систе5
174

мы от нормы, вызванную некоторой мерой возмущающих внешних воздействий.
Вданном определении устойчивость системы устанавливается, во5 первых, по отношению к ограниченному числу выбранных свойств, которые мы будем называть заданными, и, во5вторых, по отношению
кряду конкретных возмущающих воздействий среды. В этом случае об устойчивости систем можно судить по мере отклонения от нормы заданных свойств, вызванных возмущающей мерой внешнего воз5 действия. Такой подход позволяет в основу количественных оценок устойчивости положить сопоставление оценок, характеризующих, с одной стороны, меру возмущающих воздействий и, с другой – выз5 ванную меру отклонений заданных свойств от нормы. При этом возникает потребность установления не просто меры отклонения заданных свойств системы, а допустимой меры отклонений, обус5 ловленных возмущениями.
Действительно, при некоторой мере отклонений система претер5 певает необратимые изменения свойств и либо изменяет свое лицо, либо перестает быть системой. Поэтому область устойчивости конк5 ретной экосистемы определяется некоторой мерой отклонений, ко5 торые со временем могут быть ликвидированы самой системой, и она оказывается способной вернуться к исходному состоянию «нор5 мы» благодаря гомеостатическим механизмам. Эта ликвидируемая системой мера отклонений и есть допустимая мера, и ее следует считать мерой устойчивости всей системы в целом.
Взаключение этого раздела нам казалось важным обратить вни5 мание на плодотворную идею связи допустимой меры отклонений заданных свойств системы с мерой гомеостаза. Сама идея подобной связи весьма конструктивна, поскольку мера гомеостаза определена выше достаточно однозначно, а вопрос о допустимой мере отклоне5 ний (в отрыве от идеи связи с мерой гомеостаза) немедленно приоб5 ретает остроту и неясность вопроса о «норме» и «патологии». Шутливое замечание физиолога В.Г. Скребицкого, что «норма есть отсутствие явной патологии», весьма метко определяет беспомощность усилий в решении «чисто медицинского» аспекта этого вопроса. Поэтому пред5 ставляется весьма логичным специальное рассмотрение вопросов вза5 имоотношения показателей устойчивости и гомеостаза.
4. УСТОЙЧИВОСТЬ И ГОМЕОСТАЗ
Возмущающие факторы могут образовать совокупность оценок с
(9)
Возникает вопрос: в каком соподчинении находятся Sф, Sс и Sв, или, точнее  ?
?
175

Рассмотрим два аналитических примера.
Первый пример. Рассмотрим монокультуру водорослей, на кото5 рую воздействуют свет и температура. Свет индуцирует синтез био5 массы в виде возрастающего числа особей, температура регулирует скорость нарастания. Таким образом, вплоть до порога светового
насыщения показатели асвета, афотосинтеза, ачисла особей изменяются симбат5 но, и это значит, что такая система не обладает экологическим гоме5
остазом, и изменения интенсивности света целиком определяют меру отклонений в системе «водоросли – питательная среда».
Второй пример. Пусть две культуры водорослей различаются порогом светового насыщения и термофильностью. Свет индуциру5 ет синтез, но растут водоросли по5разному, т.е. численность изме5 няется с различной скоростью, вследствие чего изменяется их относительное обилие.
При этом скорость образования суммарной биомассы водорослей может быть постоянной даже при варьировании интенсивности ос5 вещения и температуры за счет изменения ростовых характеристик отдельных культур. В итоге изменяется удельная биомасса каждой культуры, что означает наличие структурной перестройки такой си5 стемы и свидетельствует о наличии экологического гомеостаза. При этом оценка гомеостаза G указывает на мощность системы, с кото5 рой она стремится к равновесию с внешними факторами среды. Из примера видно, что чем энергичнее перестраивается система, тем выше ее гомеостаз.
Для природных возмущающих начал, например, необычно высо5 ких концентраций элементов, может оказаться достаточным для оценки гомеостаза сопоставление показателей биомассы (В) и про5 дуктивности (Р*). Для того чтобы оценить гомеостаз при воздей5 ствии необычных «новых» загрязнителей, к показателям В и Р* должны быть добавлены в первую очередь те характеристики сис5 тем, Которые оказываются вовлеченными в морфологический или биохимический контакт с загрязнителями.
Возникает еще один кардинальной важности по возможным по5 следствиям вопрос: могут ли при сильном гомеостазе Sф оказаться более устойчивыми, чем Sв? Иными словами, встает вопрос об инва5 риантных свойствах живых систем, находящихся в пределах нор5 мы, когда достигнутое равновесие Sв  Sф заменяется на Sв<Sф. Нам кажется, что случай Sв<Sф, невозможен для отдельных характерис5 тик системы, связанных причинно5следственным соподчинением. Примером такого соподчинения могут быть свет и фотосинтез (све5 товые реакции), температура и фотосинтез (темновые реакции), наконец, «хищник – жертва».
Sф заменяется на Sв<Sф. Нам кажется, что случай Sв<Sф, невозможен для отдельных характерис5 тик системы, связанных причинно5следственным соподчинением. Примером такого соподчинения могут быть свет и фотосинтез (све5 товые реакции), температура и фотосинтез (темновые реакции), наконец, «хищник – жертва».
Оценки этих показателей должны быть соизмеримы по абсолют5 ным значениям Si(в)  Sj(отклик), а показатели Sв, Sc и Sф – лишь сред5
Sj(отклик), а показатели Sв, Sc и Sф – лишь сред5
176

ние отдельных оценок Si. Вследствие этого вряд ли полезно подни5 мать вопрос об инвариантности отдельных показателей систем. Они, по5видимому, не могут быть таковыми. Но первоначально вопрос был сформулирован правильно, так как это был вопрос об инвари5 антных свойствах систем, т.е. свойствах, обусловленных совокуп! ным действием комбинации изменяющихся показателей.
Инвариантные свойства биосистем а5типа известны (например, поддержание температуры тела постоянной у теплокровных живот5 ных, сохранение облика а5систем и др.). Поэтому, строго говоря, вопрос должен быть несколько сужен: обладают ли инвариантными свойствами r5системы?
Нам кажется, что предпринятая нами попытка распространения гомеостаза на системы «с плохой структурой» тем самым предопре5 деляет и положительный ответ на поставленный вопрос.
5. ИНВАРИАНТЫ КАК ПОКАЗАТЕЛИ УСТОЙЧИВОСТИ БИОСИСТЕМ
Из непостоянства отдельных переменных (оценки Si) не следует, разумеется, вывод о неустойчивости систем. Уже обобщенное толко5 вание гомеостаза логически приводит к возможности инвариант. Прав5 да, оговоримся, что оценка в виде средней (формулы (4), (5) и (9)), строго говоря, не может быть инвариантом, так как при этом мы оцениваем некоторую среднюю устойчивость отдельных показателей, которые не способны отражать более целостные характеристики со5 стояния системы. Однако, при очень грубом приближении, когда выполняется условие 0<<G=const, простейшей формой того, что мы решаемся рассмотреть в виде инварианта, оказывается все же сред5 няя оценка стабильности функциональных показателей экосистемы.
Более типичный случай инварианта, когда рассматриваемые свой5 ства в норме вообще неизменны во времени, может быть обусловлен сложной комбинацией согласованно изменяющихся отдельных пе5 ременных. Определение «согласованно изменяющихся» подразуме5 вает строгое соблюдение правил соподчинения функций, которое достигается с помощью вегетативной нервной системы в системах с центральным управлением, вследствие чего для а5систем легче отыс5 кать инварианты, чем для r5систем.
Более того. Благодаря «диффузной структуре» r5систем можно постулировать существование инвариант «в пределах нормы», т.е. не вполне строгих инвариант, которые характеризуют нормальные, устойчивые состояния экосистем. Тогда при определенных услови5 ях, которые должны быть уточнены, наличие таких «размазанных» в пределах нормы инвариантных свойств можно считать показате5 лями устойчивости экосистемы в целом.
177

Вэтом случае наблюдаемые отклонения заданных свойств за пределы нормы инвариантности могут служить сигналом выхода системы из нормы и мерой отклонений систем в целом.
Тем самым мы приходим к более целостному толкованию устой5 чивости (хотя и менее конструктивному) как поддержанию во вре5 мени инвариантных свойств благодаря комбинированному, упорядо5 ченному кооперированию непрерывно изменяющихся отдельных пе5 ременных системы.
Необходимые условия проявления инвариантных свойств биоси5 стем: 1) замкнутый характер круговорота элементов косной среды в экосистемах; 2) постоянство мгновенной скорости изменения энтро5 пии в системе.
Всоответствии с этими условиями инвариантами экологических систем могут быть: а) постоянство соотношений реакций синтеза и распада органического вещества в системе (в единицах энергии); б) постоянство соотношений скорости движения отдельных элемен5 тов через звенья трофической цепи или даже скорости отдельных элементов; в) постоянство соотношений негэнтропии, образующей5 ся на различных трофических уровнях системы.
Однако, предсказываемые на основании общих соображений пере5 численные инварианты экосистемы трудны для практического при5 менения, поскольку определение их значений требует большой пред5 варительной экспериментальной работы, а оба необходимых условия практически никогда не выполняются или выполняются не строго. В силу этих причин конкретными обобщенными показателями, которые легко измерять, могут быть: а) в пределах одного трофического звена показатель ВР* (Федоров, 1972), где В – биомасса, а Р* – скорость ее образования; б) в пределах соседних трофических звеньев показатель ВхВж, отражающий взаимодействие хищника (Вх) и жертвы (Вж).
На основании оценок ВР* и ВхВж могут быть сопоставлены урав5 нения, при конструировании которых в качестве исходных предпо5 сылок могут оказаться полезными следующие биологически правдо5 подобные допущения:
(10)
(11)
(12)
(13)
178

Важно подчеркнуть, что показатели Р*В и ВхВж не являются инва5 риантами системы в полном смысле этого слова. Можно лишь утвер5 ждать, что они в неявной форме отражают движение энергии в системе и, вероятно, могут быть использованы. Поэтому более корректно име5 новать их (впрочем, как и образующие их показатели биомассы и продуктивности) вариантами системы, постулируя, что усложнение ва5 риант, оправданное логическими предпосылками, будет асимптотичес5 ки приближать их к теоретически возможным инвариантами экосис5 темы. Логически оправданным примером такого усложнения вариант
могут оказаться показатели ВР*D и |
, где D – информаци5 |
онный индекс разнообразия (McArthur, 1955), а W – число видов.
6. ИЗМЕРЕНИЕ УСТОЙЧИВОСТИ ЭКОСИСТЕМ
После того как были определены и конкретизированы основные понятия, входящие в определение устойчивости экосистемы, можно перейти к обсуждению приемов оценки ее общей устойчивости. Ра5 зумеется, могут быть придуманы разнообразные и многочисленные способы оценок такого рода. Ниже предлагается несколько спосо5 бов такой оценки с помощью коэффициента устойчивости, который мы будем называть 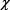 .
.
А. Канонический способ. Если мы возмущающему началу также при5пишем возможность отклонения от нормы, то мерой возмущаю5 щего действия окажется именно эта мера отклонения независимой переменной от нормы, т.е.
(14)
Тогда и результат воздействия ли отклик системы по заданным свойствам на воздействие  можно выразить относительным от5 клонением от нормы зависимой переменной, т.е.
можно выразить относительным от5 клонением от нормы зависимой переменной, т.е.
(15)
Отношение этих величин может быть названо коэффициентом устойчивости системы:
(16)
179

В норме экосистемы 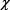 будет равен 0 при сохранении «нормы» воздействия внешних факторов, так и при поддержании механизма5 ми гомеостаза инвариантных свойств системы.
будет равен 0 при сохранении «нормы» воздействия внешних факторов, так и при поддержании механизма5 ми гомеостаза инвариантных свойств системы.
Б. Выборочный способ оценки коэффициента устойчивости c ос5 нован на учете неравномерности распределения оценок воздействую5 щего начала и регистрирующего отклика системы (формула (3)).
При этом предполагается, что показатель стабильности  изме5
изме5
ряемых переменных и рассчитанных свойств системы (вариант) ока5 зывается пригодной мерой оценки неоднородности в сопоставляемых совокупностях отдельных переменных. Иными словами, подразуме5
вается неявное тождество способа оценок  и выражения (
и выражения ( ).
).
Этот способ избавляет от необходимости кропотливой работы по оценке нормы и меры допустимых отклонений в экосистеме. Тогда
|
(17) |
где |
показатель неоднородности распределения оценок пере5 |
менных, рассчитанных по формуле (3). Важно отметить, что откли5 ком системы в этом случае служит оценка одной из вариант, которой мы приписываем решающее значение при определении состояния системы (например, оценка по ВР*D). При этом в качестве возму5 щающего начала также используется оценка стабильности только одной из независимых переменных, действию которой мы приписы5 ваем отклик системы.
В. Вариантом выборочного способа является оценка коэффици5 ента устойчивости по средним значениям совокупности оценок Si , относящихся к воздействующим началам и «откликам системы»:
(18)
Назовем эту разновидность рекогносцировочным способом, при5 мени5мым при изучении комбинированного воздействия возмущаю5 щих начал на экосистему.
Г. Способ трех средних использует для оценки коэффициента устойчивости средние значения стабильностей, определенных лю5 бым из способов, относящиеся к оценкам возмущающего начала 
и состояния системы –  и
и  .
.
180