

Изменения в природных биологических системах - Федоров В.Д
..pdfтичной, так как неизвестно, какие именно связи надо учитывать и как измерять силу этих связей между индивидуумами.
13) Стиль последней фразы заставляет вспомнить выступления ру! ководителей партии и правительства на очередном пленуме ЦК. Но, как в этих выступлениях, так и здесь, — «гладко было на бумаге...», а на практике за прошедшие 30 лет мы не очень!то продвинулись в анали! зе многообразия взаимодействий, а уж с прогнозом поведения сложных систем и подавно дело обстоит весьма печально. Поневоле задумыва! ешься о том, что на пути редукционизма нам не стоит рассчитывать на серьезные успехи даже в условиях всеобщей компьютеризации.
161

НОВЫЙ ПОКАЗАТЕЛЬ НЕОДНОРОДНОСТИ СТРУКТУРЫ СООБЩЕСТВА
В гидробиологии и экологии часто возникает потребность коли5 чественно охарактеризовать распределение организмов (или любых признаков, относящихся к сообществу) в пространстве биотопа. Пред5 лагаем один из таких способов.
Разделим пространство биотопа (например, столб воды под 1м2) на k любых частей, каждая из которых содержит 1/k всех особей. Тогда отрезок пространства L, равный глубине водоема в точке от5 бора проб, разобьется на k отрезков:
L = l1 + l2 + ... + lk
Легко убедиться, что максимальным окажется произведение длин отрезков  при l1 = l2 = ... =lk, соответствующее равномер5 ному распределению организмов в пространстве. Это произведение может служить некоторой мерой однородности (или неоднороднос5 ти) распределения признака на участке L. Для того чтобы сделать такой показатель независимым от абсолютной величины отрезка L, разделим каждый из сомножителей на среднее значение
при l1 = l2 = ... =lk, соответствующее равномер5 ному распределению организмов в пространстве. Это произведение может служить некоторой мерой однородности (или неоднороднос5 ти) распределения признака на участке L. Для того чтобы сделать такой показатель независимым от абсолютной величины отрезка L, разделим каждый из сомножителей на среднее значение
Тогда мера неоднородности U может быть записана как относи5 тельная величина:
(1)
В этом случае показатель неоднородности характеризуется произ5 ведением длин отрезков, относящихся к пространствам с одинако5 вым числом особей. При максимальной однородности распределения
особей, когда  , показатель U оказывается равным 1, тогда как
, показатель U оказывается равным 1, тогда как
при всех li  l его значения будут отклоняться в сторону нуля. Вместе с тем, другим выражением неоднородности может слу5
l его значения будут отклоняться в сторону нуля. Вместе с тем, другим выражением неоднородности может слу5
жить произведение численности особей — Ni, (или каких5либо мер признака) в точках, равнорасположенных на отрезке L. Тогда по аналогии с формулой 1 показатель U может быть записан как
(2)
В зависимости от характера сбора данных может быть применен тот или иной из предложенных показателей неоднородности. Так, например, учитывая особенности современной техники сбора зоо5 планктона (отлов не в отдельных дискретных точках, а в ограни5
162

ченном числе горизонтов столба воды), для характеристики неодно5 родности распределения особей под 1 м2, вероятно, более предпоч5 тительным оказывается показатель, рассчитываемый по формуле 1, поскольку длины отрезков li могут быть легко вычислены по Снеде5 кору (1961, стр.120) или получены графическим путем, если по оси ординат отложены накопленные частоты признака, а по оси абс5 цисс — интервалы пространства или времени. Так, для зоопланкто5 на удобно рассчитывать так называемые квартили, каждая из которых содержит 25% всех особей, находящихся под 1м2 поверхности.
Показатель неоднородности прост в употреблении, биологически легко осмысливается и зависит от выбора числа сомножителей (k или n). Последнее обстоятельство вынуждает принять необходимое ограничение: по индексу неоднородности сравнивать следует сооб5 щества только с одинаковым значением n, т.е. число точек и рассто5 яние между ними должны быть сходными. Аналогичные требования предъявляются и по отношению k, с той, однако, разницей, что выбор доли особей (т.е. 1/k) автоматически предопределяет число облавливаемых горизонтов.
Для сопоставления оценок неоднородности, полученных с раз5 личными k, полезно ослабить зависимость U от числа сомножите5 лей. Простейшая операция «по ослаблению» связана с приведением оценки к новому общему значению, что можно достичь извлечением корня степени, равной числу сомножителей. Тогда формула 1 при5 обретает следующий вид:
(3)
Аналогичным образом может быть преобразована формула 2:
(4)
Однако, как видно из таблицы, произвол в разбиении L на n от5 резков при пользовании формулой 2 может послужить источником дополнительных ошибок в оценке U. Действительно, деление отрез5 ка L с учетом распределения накопленных частот исключает откры5 тие неоднородности, связанное со случайным попаданием точки на моду, соответствующую наибольшей численности в анализируемой совокупности. Это дает нам основание считать оценки неоднороднос5 ти по формулам 1 и 3 более правдоподобными, чем по формулам 2 и 4. Поэтому расчет квартилей, децилей, персентилей, промиллей и т.п. позволяет произвести дробление ряда наблюдений L на k час5
163

164
Оценка U совокупности наблюдений, заданной рядом, по произвольно сделанным выборкам
Ка |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
u |
U |
вы5 |
п |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
форм. |
форм. |
||
борки |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
24 |
2 |
2 |
2 |
3 |
5 |
6 |
6 |
5 |
4 |
2 |
1 |
1 |
1 |
1 |
2 |
3 |
4 |
4 |
3 |
2 |
3 |
6 |
5 |
1 |
0,017 |
0,844 |
1 |
12 |
|
2 |
|
3 |
|
6 |
|
5 |
|
2 |
|
1 |
|
1 |
|
3 |
|
4 |
|
2 |
|
6 |
|
1 |
0,098 |
0,824 |
2 |
12 |
2 |
|
|
|
5 |
|
6 |
|
4 |
|
1 |
|
1 |
|
2 |
|
4 |
|
3 |
|
3 |
|
5 |
|
0,170 |
0,863 |
3 |
8 |
|
2 |
|
|
5 |
|
|
5 |
|
2 |
1 |
|
|
1 |
|
|
4 |
|
|
2 |
|
6 |
5 |
|
0,220 |
0,826 |
4 |
8 |
2 |
|
|
3 |
|
|
6 |
|
|
|
|
1 |
|
|
3 |
|
|
3 |
|
|
|
|
0,312 |
0,865 |
||
5 |
6 |
|
2 |
|
|
|
6 |
|
|
|
2 |
|
|
|
1 |
|
|
|
4 |
|
|
|
6 |
|
|
0,313 |
0,824 |
6 |
6 |
2 |
|
|
|
5 |
|
|
|
4 |
|
|
|
1 |
|
|
|
4 |
|
|
|
3 |
|
|
|
0,476 |
0,884 |
7 |
4 |
|
2 |
|
|
|
|
|
5 |
|
|
|
|
|
1 |
|
|
|
|
3 |
2 |
|
|
|
|
0,513 |
0,846 |
8 |
4 |
2 |
|
|
|
|
|
6 |
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
0,444 |
0,816 |
9 |
3 |
|
2 |
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
0,895 |
0,964 |
10 |
3 |
2 |
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
0,866 |
0,953 |
11 |
3 |
2 |
|
2 |
|
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0,741 |
0,905 |
12 |
4 |
2 |
|
2 |
|
5 |
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0,607 |
0,883 |
13 |
5 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0,606 |
0,905 |
14 |
6 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
0,352 |
0,840 |
15 |
7 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
; |
|
|
|
|
|
|
|
|
|
|
|
0,219 |
0,805 |
16 |
8 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
; |
|
2 |
|
|
|
|
|
|
|
|
|
0,206 |
0,821 |
17 |
9 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
|
|
2 |
|
4 |
|
|
|
|
|
|
|
0,195 |
0,834 |
18 |
10 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
1 |
|
2 |
|
4 |
|
3 |
|
|
|
|
|
0,195 |
0,849 |
19 |
11 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
1 |
|
2 |
|
4 |
|
3 |
|
3 |
|
|
|
0,195 |
0,862 |
20 |
12 |
2 |
|
2 |
|
5 |
|
6 |
|
4 |
|
1 |
|
1 |
|
2 |
|
4 |
|
3 |
|
3 |
|
5 |
|
0,170 |
0,863 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

тей, каждая из которых содержит 1/k5тую часть особей (признака). При этом, независимо от формы распределения совокупности, путем перемножения длин отрезков li мы получаем некоторое приближен5 ное описание выборки. Так, таблица свидетельствует, что при харак5 теристике совокупности, представленной рядом L (первая строка таблицы), оценка U по формуле 3 дает близкие значения при разби5 ении L на квартили, пентили, октили и децили (0,966; 0,938; 0,914; 0,918 соответственно). В то же время оценки U, произведенные по формуле 4, обнаруживают несколько больший разброс (таблица), в чем можно убедиться, сравнив коэффициенты вариации (v = 2,5% при оценке U по формуле 3 и v = 4,8% при оценке по формуле 4).
С помощью U может быть охарактеризована неоднородность любого признака по выбранному трансекту, а также оценена мера устойчивости признака, изменяющегося по времени (при замене интервалов пространства на интервалы времени).
The new index of irregularityof the structure of a community
Summary
A new index of irregularity for the structure of a community is proposed:
where k is the number of biotope parts, every of which contains 1/k of all individuals of the population under investigation, and li is the section length of space characteristic of every part. By U the uneveness of every parameter distribution at chosen transect can be characterized and also the measure of stability for time dependent parameter (when space intervals are considered instead of time intervals).
Л и т е р а т у р а :
Снедекор Дж. У. Статистические методы в применении к исследовани5 ям в сельском хозяйстве и биологии. М., Сельхозгиз, 1961.
Вестник МГУ, биол., № 2, 1973.
165

УСТОЙЧИВОСТЬ ЭКОЛОГИЧЕСКИХ СИСТЕМ И
ЕЕИЗМЕРЕНИЕ
1.ОБЩИЕ ОСОБЕННОСТИ ОРГАНИЗАЦИИ БИОЛОГИЧЕСКИХ СИСТЕМ
Совокупность взаимодействующих элементов образует систему (Bertalanffy, 1952). Всякая система организована в том смысле, что ее поведение подчинено правилам, определяющим способы и особен5 ности взаимодействия образующих ее элементов. Благодаря этому в любой системе имеет место упорядоченность во времени и простран5 стве отдельных переменных и разнообразных свойств системы, ха5 рактеризующих ее состояние. Одна из основных форм упорядо5 ченности – соподчинение групп элементов и связанных с ними свойств, вследствие чего система обладает иерархией структуры, что позволя5 ет говорить об уровне ее организации. Системный подход не конкре5 тизирует понятие «элемент», вследствие чего выбор элемента зависит от уровня организации системы. Известная свобода в выборе того, что следует считать элементом системы, зависит всякий раз от свойств системы, являющихся предметом исследования. Поэтому в каждом конкретном случае элементы системы следует выбирать с таким рас5 четом, чтобы влиянием их внутренних свойств на изучаемые свой5 ства системы можно было пренебречь.
Так, например, для биологических систем элементами могут быть макромолекулы (при рассмотрении какого5либо типа клеток в каче5 стве системы), клетки (при рассмотрении в качестве системы кле5 точной ткани), ткани (по отношению к органу), органы (по отноше5 нию к целому организму) и, наконец, особи или популяции при системном изучении сообщества или биоценоза).
Главная особенность биологических систем – сложность, кото5 рая предопределяет основные черты их организации. Принято счи5 тать, что все биологические системы «плохо организованы» в том смысле, что у них не удается выделить для исследования в чистом виде отдельные явления и процессы одной физической природы (На5 лимов, 1971). Действительно, в таких системах «с плохой структу5 рой», каковыми являются биологические системы, действует множест5 во физически разнородных факторов, что обусловливает объектив5 ные трудности в изучении состояния и поведения систем. Невоз5 можность изучить все переменные системы такой повышенной сложности неизбежно привносит субъективный момент при отборе немногих из них, которые оказываются с точки зрения специалиста наиболее важными для понимания сути исследуемого явления. Ап5 риорное ранжирование переменных систем по важности (произве5 денное сознательно или бессознательно) оказывается поэтому
166

необходимым этапом, предшествующим исследованию биологичес5 ких систем любой сложности (Федоров, 1972).
Вынужденная необходимость оперировать ограниченным числом переменных при изучении интегральных свойств очень сложных систем не позволяет установить простых причинно5следственных форм зависимости между разнообразными свойствами биологичес5 ких систем. Это породило в биологии точку зрения, согласно кото5 рой целое оказывается больше составляющих его частей, вследствие чего свойства целого не могут быть эффективно исследованы после разделения целого на части на основе изучения свойств последних. В результате такого подхода появилась мысль о существовании осо5 бых биологических законов, которые не могут быть объяснены с позиций строгих положений таких наук, как физика и химия.
Вероятно, подобный взгляд на вещи слишком тенденциозен, од5 нако, он как в зеркале отражает положение дел, существующее се5 годня в биологии в период ее становления как количественной науки. Вместе с тем, эта точка зрения достаточно плодотворна в настоящее время, так как выдвигает на повестку дня исследование интеграль5 ных, целостных свойств биологических систем, которые отражают лицо систем – их облик, судьбу и соподчинение взаимодействую5 щих элементов. В связи с этим можно дать определение системы, подчеркивающее ее целостные свойства.
Определим биологическую систему как способный к образова5 нию негэнтропии объект, целостные свойства которого есть резуль5 тат взаимодействия образующих его частей, и выделим в качестве основных следующие целостные свойства биологических систем:
1)сложность (определяется разнообразием элементов и связей между ними, а также напряженностью связей в системе);
2)автономность (определяет особенности положения системы на правах подсистемы в пределах более крупной системы);
3)надежность (определяет вероятность осуществления процес5 сов в системе и поддержания во времени постоянства ее свойств);
4)устойчивость (определяет способность биосистем противо5 стоять внешним возмущающим воздействиям при сохранении «лица» системы, зависящего от ее предыстории).
Несомненно, что для конкретных биологических систем ранжирова5 ние целостных свойств по важности может варьировать в зависимости от того, какими особенностями обладает система и какие задачи решает
вконкретном случае исследователь. Вместе с тем существуют объек5 тивные основания для разделения биологических систем на три группы
взависимости от различий в регуляции их поведения (Федоров, 1972).
1.Биологические системы а5типа с центральным управлением, т.е. подсистемой, регуляторная связь которой с любым элементом силь5 нее, чем связь этого элемента с любым другим произвольно взятым.
167

К этой категории относятся все животные организмы с централь5 ной и вегетативной нервной системами.
2.Биологические системы р5типа с «пассивным» управлением (Milsum, 1964), куда можно отнести растительные организмы с их гуморальной регуляцией.
3.Биологические системы r5типа со «случайным (random)» уп5 равлением, куда можно отнести все экологические системы, не не5 сущие специальной «программы» поведения в молекулах ДНК, как это имеет место в первых двух случаях.
Вполне естественно, что для биологических систем различных типов и важность перечисленных целостных свойств будет различ5 на. Так, Старостин (1967) обратил внимание на то, что системы без центрального управления менее жестко детерминированы и поэтому судьба их (устойчивость – В.Ф.) менее зависит от поражения мел5 ких участков системы. Несомненно поэтому, что для систем, облада5 ющих «программой» (а и р5тип), первостепенное значение будут иметь такие целостные свойства, как надежность и автономность, от которых в первую очередь зависит выполнение «программы». В то же время для экологических систем (r5тип) первостепенное значе5 ние будут иметь такие целостные характеристики, как сложность и устойчивость, от которых в первую очередь зависит их судьба, т.е. способность противостоять случайным «незапрограммированным» возмущениям путем перестройки структуры в направлении дости5 жения равновесия с условиями, господствующими на определенном пространстве обитания (локусе).
Настоящая работа посвящена обсуждению некоторых общих по5 ложений устойчивости экологических систем и обоснованию спосо5 бов ее измерения. Поэтому следует начать с краткого обсуждения особенностей систем r5типа, которые будут необходимы для после5 дующего изложения наших взглядов на устойчивость.
Известно, что экологическая система образована биотической и абиотической компонентами, т.е. населяющими биотоп организмами
ивнешней средой, определяющей условия существования жизни в пределах биотопа. Обе компоненты образуют некоторое единство, поскольку не только внешняя среда оказывает влияние на живую компоненту, но и, наоборот, живая компонента изменяет окружаю5 щую среду. Это выражается в том, что фотосинтезирующие организ5 мы непрерывно вовлекают элементы окружающей среды в цепь превращений, образованную цепочками организмов, взаимосвязан5 ных отношениями «хищник – жертва». Таким образом элементы дви5 гаются от организма к организму по пищевым (трофическим) цепям,
итем самым поддерживается неразрывное единство экосистемы, фор5 мирующее ее структуру, поведение и нормы реакции на окружаю5 щие воздействия. Факторы внешней среды, вовлекаемые во взаимо5
168

действие живой компонентой системы, т.е. оказывающие на нее воз5 действие и испытывающие ответное воздействие со стороны орга5 низмов, должны быть отнесены к внутрисистемным. К таким внутрисистемным факторам относятся, прежде всего, все химичес5 кие элементы, изменяющиеся в экосистеме в результате включения
впищевые цепи, которые «в норме» замкнуты, образуя биогеохи5 мические циклы, формирующие облик нашей планеты. Как прави5 ло, в отдельных локусах общий запас тех или иных элементов мало изменяется и в интервале времени, соизмеримом с жизнью данного типа системы, может считаться постоянным.
Вторую группу факторов внешней среды образуют внесистем5 ные факторы, которые определяют условия существования конк5 ретных экосистем в пределах более крупных системных образований, не испытывая при этом заметного ответного воздействия со стороны биотической компоненты. К ним следует отнести солнечную радиа5 цию, температуру, влажность и соленость. Но поскольку биотичес5 кая компонента эволюционировала под влиянием параллельного развития жизни и изменения этих факторов, то она хорошо при5 способилась к их сезонным, годичным и другим колебаниям. Вслед5 ствие этого периодические флюктуации внесистемных факторов не угрожают существованию экологических систем.
Наконец, третью группу факторов образуют возмущающие фак5 торы, к воздействию которых экосистемы мало подготовлены (или не подготовлены вовсе) предшествующим ходом эволюции видов и предысторией формирования конкретных сообществ в условиях оп5 ределенных биотопов. К их числу могут принадлежать физически разнородные факторы, но их отличительной особенностью оказыва5 ется внезапность (в геологическом масштабе времени) воздействия, вызванная необычно резким для системы изменением их интенсивно5 сти. К такому воздействию может быть отнесено резкое изменение запаса какого5либо «обычного» элемента в локусе (например, N и Р, породивших проблему эвтрофикации), или обычно редкого элемен5 та (например, соли тяжелых металлов – Hg, Pb, Cd), или, наконец, появление нового для жизни вещества, синтезированного челове5 ком (например, ДДТ, РСВ и другие пестициды).
Таким образом, воздействие необычно больших количеств при5 родных элементов наряду с воздействием малых доз новых, создан5 ных человеком веществ образует совокупность возмущающих фак5 торов, к воздействию которых экосистемы оказались не подготовле5 ны всем ходом их эволюции.
Именно интенсификацией возмущающих факторов, связанных с разнообразием форм и усилением масштабов человеческой деятель5 ности в биосфере, и объясняется та первостепенная важность, кото5 рую приобрела проблема изучения устойчивости природных систем
внастоящее время.
169

2. ИЗМЕРЕНИЕ СТАБИЛЬНОСТИ ПЕРЕМЕННЫХ
Биологическая система обладает множеством характеристик, от5 ражающих особенности ее организации. Все они изменяются во вре5 мени и поэтому определяют изменение состояния системы во времени. Все они изменяются в пространстве, поддерживая существование разнообразных градиентов, и тем самым обусловливают динамичес5 кий характер пространственной структуры системы. Такие характе5 ристики могут быть рассмотрены в качестве независимых переменных системы, и при любом изучении системы постоянно возникает по5 требность оценить их поведение во времени и (или) в пространстве. Иными словами, должны существовать способы, позволяющие оце5 нить поведение отдельных переменных системы. Для оценки измене5 ния во времени отдельной независимой переменной предлагается измерение ее стабильности (Si) в качестве оценки колебания разброса единичных значений функции вокруг среднего. Таким образом, ста5 бильность определяется, как мера непостоянства любой функции от времени. Такими оценками Si могут быть (Федоров, Соколова, 1972) средние значения отклонения амплитуды, 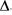 (1), или квадрата этого отклонения,
(1), или квадрата этого отклонения,  (2), переменной относительно ее среднего значения:
(2), переменной относительно ее среднего значения:
(1)
(2)
где хi – значение измеряемой переменной в момент измерения, а n – число измерений при  и
и
Другим способом оценки Si может служить предложенный нами (Федоров, 1973) показатель неоднородности распределения признака во времени и (или) в пространстве, который является оценкой соот5 ношения среднего геометрического к среднему арифметическому значений переменной –  (3):
(3):
(3)
170