

Изменения в природных биологических системах - Федоров В.Д
..pdf(х3), кремния (х4) и аммиачного азота (х5). За нижний уровень (5) в плане принимали природную концентрацию элементов в момент по5 становки опытов. Верхний (+) – получали добавкой в опытные склян5 ки соответствующей соли элемента из расчета:
для нитратного азота |
– 80 мкг N/л в виде NaNO3 |
|
>> фосфора |
– 20 мкг Р/л в виде NaHPO4 |
|
>> железа |
– 100 |
мкг Fe/л в виде FeSO4 |
>> кремния |
– 500 |
мкг Si/k в виде SiO2 |
>> аммиачного азота |
– 10 мкг N/л в виде NH4Cl |
|
В 1968 г. поставлено 14 серий опытов по плану ПФЭ 24 [2], в которых исследовалось влияние на изменение состава фитопланкто5 на нитратного азота (х1), фосфата (х2) и железа (х3) при двух уров5 нях освещенности (х4). Величины добавок первых трех солей равнялись 100 мкг N/л, 20 мкг Р/л и 50 мкг Fe/л. За верхний уровень освещенности принята естественная освещенность в повер5 хностном (0,1–0,3 м) горизонте. Двукратного ослабления освещен5 ности (нижний уровень) в эксперименте достигали экранированием соответствующих склянок несколькими слоями марли; ослабление радиации контролировали пиранометром Янишевского.
Каждому варианту опытов в матрицах планирования соответство5 вала опытная склянка с 1,5 л морской воды, отобрано в день поста5 новки опытов с поверхностного (0,2–0,3 м) горизонта. Воду отфильт5 ровывали через мельничный газ № 23 для удаления крупного зоо5 планктона, после чего в каждую склянку добавляли элемент в соот5 ветствии с одной из комбинаций факторов в матрице эксперимента.
Подготовленные таким образом склянки с пробами экспонирова5 ли in situ на глубине 0,2–0,3 м течение пяти (1967 г.) и четырех (1968 г.) суток, затем фиксировали раствором люголя, и после от5 стаивания и концентрирования учитывали под микроскопом в каме5 ре типа Нажотта видовой состав фитопланктона в соответствии с нашими рекомендациями [1].
Данные подсчета клеток в отдельных вариантах каждого опыта для каждого вида использовали при расчете показателя биотическо5 го разнообразия в пробе, соответствующей одному из вариантов в матрицах ДФЭ 25–1 и ПФЭ 24 по формуле:
Полученные значения D для каждой серии экспериментов обра5 ботаны по Ятсу [2]; выделенные согласно Дэниелу [2] значимые эффекты использовали для получения уравнений регрессии, связы5 вающих величины разнообразия – D с изменением в эксперименте уровней изучаемых факторов.
131
Всего получено семь уравнений регрессии для серий опытов 1967 г. (индекс при «D» указывает порядковый номер серии):
D2 = 0,618 – 0,074x2x4x5;
D9 = 0,406 + 0,038x1– 0,049x2– 0,040x1x2 – 0,037x2x3x5; D10 = 0,425 – 0,037x2 – 0,029x2x3 – 0,036x1x4 – 0,023x2x3x4; D11 = 0,378 + 0,103x2 – 0,101x2x3x5 – 0,076x3 – 0,066x2x5; D12 = 0,510 – 0,055x1x2;
D13 = 0,344 – 0,113x2;
D14 = 0,443 – 0,053x4 – 0,077x2x5 – 0,046x1x5 – 0,066x5; и восемь урaвнений для опытов 1968 г.
D3 = 0,540 – 0,097x2;
D5 = 0,556 – 0,107x1x2x4;
D6 = 0,651 – 0,072x4 – 0,083x1x3x4;
D7 = 0,635 – 0,047x2 – 0,040x1x2x4;
D8 = 0,360 + 0,049x1 – 0,063x3 – 0,047x1x2x3;
D9 = 0,427 – 0,048x2 – 0,051x1x4 – 0,044x2x4; D13 = 0,331 – 0,067x4;
D14 = 0,466 – 0,062x4 – 0,046x1x2x4.
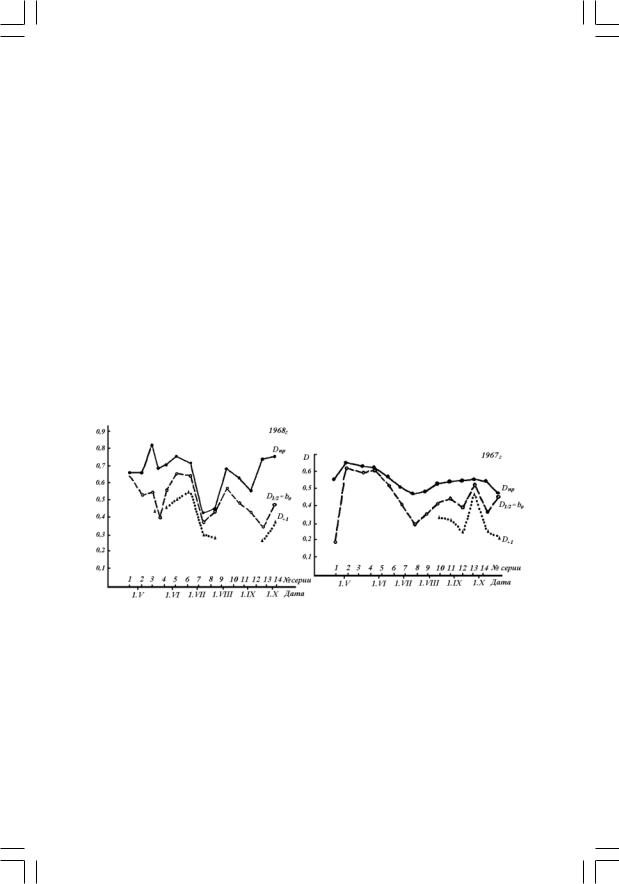
Сезонные изменения показателя видового разнообразия природного фитопланктона Dmp после постановки обогатительных экспериментов (D1/2, D+1)
В семи сериях опытов 1967 г. и шести – 1968 г. выделить значи5 мые эффекты не удалось. Это свидетельствовало, что в период их постановки связь изменения разнообразия фитопланктонного сооб5 щества с колебанием в эксперименте концентрации отдельных эле5 ментов достоверно не могла быть оценена количественно. Вместе с тем, даже в таких опытах суммарный эффект влияния изменений условий на разнообразие можно охарактеризовать средним арифме5
132

тическим всех значений Р, полученных во всех вариантах каждой серии опытов (тем же способом получены величины b0 во всех при5 веденных выше уравнениях регрессии). Эти величины соответству5 ют мыслимому опыту в центре эксперимента, т.к. изменения D при этом вызваны половинным варьированием изучаемых в опыте фак5 торов  Значения
Значения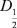 были рассчитаны для всех 28 реализо5
были рассчитаны для всех 28 реализо5
лизованных серий.
С помощью полученных уравнений найдены величины разнообра5 зия (D+1) при максимальном для опыта изменении условий мине5 ральной обеспеченности (когда факторы, входящие в соответствующее опыту равенство, находятся на верхнем уровне, т.е. подставляются в уравнение как кодированные переменные, равные +1) и сохранении природной освещенности (для 1968 г. х4 = +1).
Помимо обработки опытных данных, в дни окончания экспери5 ментов с поверхностного горизонта водоема отбирали пробы фито5 планктона, для которых также рассчитывали индекс разнообразия природного сообщества – Dпр. Эти значения использовали для срав5 нения с величинами  и D+1 ,полученными в результате обработки
и D+1 ,полученными в результате обработки
данных факторных экспериментов.
На рисунке показаны изменения видового разнообразия фитоплан5 ктонного сообщества в течение вегетационных сезонов 1967–1968 гг. Кривые характеризуют сезонные изменения разнообразия в природ5 ных условиях (Dпр), то же по данным опытов в центре экспери5 мента ( )и, наконец, воздействие в отдельные периоды сезона на
)и, наконец, воздействие в отдельные периоды сезона на
разнообразие условий, максимально измененных постановкой экс5 перимента (D+1 и точки, обозначенные  ).
).
Приведенные данные (см. рисунок) убеждают, что в преоблада5 ющем числе случаев изменение условий минерального питания умень5 шает видовое разнообразие сообщества, поскольку для каждой серии опытов независимо от периода вегетационного сезона оказывалось справедливым неравенство:
Любопытно, что снижение освещенности в опытах 1968 г. (при х4= – 1) оказывает на величину индекса D влияние, как правило, противоположное действию минеральных компонентов. Это хорошо согласуется с более общим заключением, согласно которому не вся5 кие изменения окружающей среды приводят к уменьшению разнооб5 разия планктонного сообщества, а лишь те из них, в которых ликвидируется или ослабляется ограничивающее развитие фитопланк5 тона действие условий окружающей среды. Именно поэтому при ста5
133

билизации внешних условий, когда все процессы в системе сбаланси5 рованы и, следовательно, ограничены интенсивностью прихода и рас5 хода энергии и веществ, разнообразие сообщества повышается. Это последнее находится в хорошем соответствии с отмеченным выше ис5 ходным фактом, согласно которому районы с более «суровыми» усло5 виями среды и отчетливо выраженной картиной сезонных изменений отличаются более низким разнообразием живых форм, чем районы низких широт.
Л и т е р а т у р а :
Кольцова Т.И и др. Гидробиол. Ж., 7. 1071. № 3.
Максимов В.Н., Федоров В.Д. Применение методов математического планирования экспериментов при отыскании оптимальных условий куль5 тивирования микроорганизмов. М., МГУ, 1969.
Федоров В.Д. Научн. докл. высшей школы, серия биол. н. 1970. № 2.
Федоров В.Д., Семин В.А. Океанология, 1970. № 2.
Федоров В.Д., Белая Т.И., Максимов В.Н. Известия АН СССР, се5 рия биол. н., 1970. № 3.
Margalef R. Gen. syst., 1957. № 3.
Pielou E.C., Arnason A.N. Science, 1966. 151.
Connection of the phytoplankton species diversity with changes in the mineral nutrition conditions
Summary
In concentration experiments in situ, the influence of phosphorus, iron, silicon, nitrate and ammonia nitrogen on the phytoplanktic association species diversity have been investigated.
Fifteen regression equations have been involved in order to link the D index with experimental changes at the level of factors investigated. A conclusion is made, that environmental changes limiting or excluding the action of factors pressing the phytoplankton development, lead to a decrease of D value.
Гидроб. Ж., № 3, 1972.
1) Если рассматривать эту статью в отрыве от других публикаций В.Д. Федорова (в соавторстве с участниками экспедиций 1967 и 1968 гг.), то можно подумать, что вся эта чудовищная по объему эксперимен! тальная работа была проделана только для проверки достаточно три!
134

виального предположения об уменьшении видового разнообразия планк! тона при резком изменении абиотических условий. В действительнос! ти здесь представлен лишь один аспект из всего комплекса изучавшихся проблем, а то, что в центре внимания оказался все тот же индекс Шеннона, отражает лишь то преувеличенное значение, которое прида! вала ему в те годы «передовая научная общественность».
2)Представляю, какое изумление, смешанное с возмущением, испы! тывали современники, увидев всю эту статистическую абракадабру, тем более, что в данной статье автор не счел нужным дать хоть ка! кую!нибудь интерпретацию тем эффектам взаимодействий, которые оказались статистически значимыми. Впрочем, я бы и сейчас не стал этого делать. Подробное рассмотрение каждого из вышеприведенных уравнений регрессии могло бы составить предмет отдельной статьи, а общий вывод все равно сводился бы к тому, что влияние каждого из факторов минерального питания на видовую структуру тесно связано
суровнем других, и характер этой связи меняется по сезону.
3)Следует отдать должное изобретательности автора, который сумел!таки получить некоторые общие выводы из столь пестрого набо! ра экспериментальных данных, на первый взгляд, мало согласованных
между собой.
135

ОБ ЭКОЛОГИЧЕСКИХ НИШАХ, ЛОКУСАХ БИОТОПА И ЭВОЛЮЦИОННОМ РАЗНООБРАЗИИ ВИДОВ
Рассмотрены особенности экологических ниш микроорганизмов и обсуждены возможные следствия различной соразмерности ско5 рости изменения среды обитания (Vс) и скорости изменения эколо5 гических ниш (Vн). Вариант Vн>>Vс ближе всего соответствует микроорганизмам, вес отдельных особей которых измеряется мик5 рограммами (бактерии, водоросли). Вариант V  V соответствует мезоорганизмам, вес которых удобно выражать в граммах (насеко5 мые). Наконец, вариант V<<V соответствует макроорганизмам, вес которых удобно выражать в килограммах (позвоночные). Показа5 но, что относительное разнообразие видов (W) при реализации в природе всех трех рассмотренных вариантов должно подчиняться
V соответствует мезоорганизмам, вес которых удобно выражать в граммах (насеко5 мые). Наконец, вариант V<<V соответствует макроорганизмам, вес которых удобно выражать в килограммах (позвоночные). Показа5 но, что относительное разнообразие видов (W) при реализации в природе всех трех рассмотренных вариантов должно подчиняться
неравенству Wмакро <Wмикро<Wмезо.
Эколог, исследующий микроскопические формы жизни, вынуж5 ден мириться с ограничениями, обусловленными малыми размера5 ми объекта, которые практически делают невозможным исследование судьбы отдельных особей в сообществе. Поэтому в микробиологи5 ческих исследованиях суть методического подхода заключается в изучении особенностей превращений разнообразных субстратов, свя5 занных с активностью целых популяций. В этих условиях особо важное значение приобретает масштаб процесса, характеризующий его протяженность в пространстве и времени.
В лаборатории крупномасштабность процессов достигается со5 зданием подходящих условий, в которых реализуются те или иные физиологические потенции вида. Однако при этом мы получаем све5 дения о том, что могут делать организмы, а не что они делают в природе на самом деле. При всей грандиозности масштаба превра5 щений, осуществляемых в природе микроорганизмами, отдельные процессы, вызванные активностью популяции конкретного вида, как правило, мелкомасштабны. Наблюдаемый в природе итоговый зри5 мый результат деятельности микроорганизмов слагается из бесчис5 ленного множества локализованных, часто весьма специфичных процессов, каждый из которых связан с активностью популяции отдельного вида. Возникает вопрос: а почему, собственно, в массе своей процессы, вызываемые активностью отдельных популяций мик5 роорганизмов, мелкомасштабны? Почему величественное здание слагается из множества кирпичей, а не из ограниченного числа бе5 тонных плит внушительного размера?
Чтобы ответить на этот вопрос, полезно обратиться к представ5 лениям об экологической нише вида. Согласно современным пред5 ставлениям, фундаментальная ниша вида объединяет совокупность
136

всех точек замкнутой области в n5мерном пространстве. Внутри та5 кой области каждая точка допускает существование вида. Каждая из осей координат соответствует отдельному фактору, от которого зависит существование вида, и может быть выражена в виде одно5 мерной матрицы дискретных значений внешних факторов [2] и от5 кликов организма на их воздействие [3]. Таким образом, эколо5 гическая ниша отражает совокупность требований вида к условиям окружающей среды независимо от конкретной природы отдельных факторов, воздействующих на судьбу вида. Понятие фундаменталь5 ной ниши поэтому шире пищевых потребностей вида. Оно включает также установление зависимости существования вида от условий освещения, температуры, солености, активной кислотности и т.д. в которых вид поддерживает себя неопределенно долгое время.
Итак, фундаментальная ниша в указанном выше смысле опреде5 ляется физиологическими потенциями вида, наследственно закреп5 ленными его генотипом. Реализация этих потенций зависит от степени соответствия существующих условий конкретного биотопа потреб5 ностям вида и поэтому характеризует фенотип вида. Следователь5 но, фенотип есть продукт взаимодействия генотипа с внешней средой.
Экологические ниши микроскопически малых организмов обна5 руживают, по крайней мере, две особенности, отличающие их от ниш более высоко организованных организмов. Во5первых, они срав5 нительно просты, вследствие чего в лаборатории обычно удается воссоздать условия, удовлетворяющие требованиям отдельных ви5 дов. Во5вторых, градиентная протяженность относительно каждой из определяющих нишу координат сравнительно невелика (при вы5 ражении ее в абсолютных единицах принятых измерений) для боль5 шинства пищевых факторов и факторов внешней среды (например, существования в узкой зоне рН, концентрации доступных питатель5 ных веществ и т.д.).
При проекции ниши на биотоп масштаб процесса, осуществляемо5 го видом, будет целиком определяться реальной протяженностью ус5 ловий, которые соответствуют данной нише. Такой участок биотопа, в который «укладываются» точки пространства ниши и, следователь5 но, в котором вид способен поддерживать себя неограниченно долгое время, будем называть локусом. Поэтому одной и той же нише может соответствовать участок биотопа различного размера по площади или объему. Таким образом, вопрос о масштабности процессов, определя5 емых размером локуса, отвечающего требованиям вида, есть вопрос о соответствии структуры экологической ниши структуре биотопа.
Любой природный биотоп структурирован. Его структура опре5 деляется наличием всевозможного рода градиентов, отражающих изменение однородности его свойств в четырехмерном (четвертое измерение – время) пространстве. Поэтому среду обитания, кото5
137

Рис. 1. Два возможных типа (А и Б) возрастания числа фокусов при «дроблении» макроструктурированного биотопа
рую можно характеризовать наличием только микроградиентов, сле5 дует считать структурированной по отношению только к микроте5 лам и микропроцессам. Вместе с тем по отношению к макротелам такая среда может считаться относительно изотропной. Хорошим при5 мером относительной структурированности среды обитания являют5 ся поверхностные горизонты водоема, относительно однородные для рыб и весьма неоднородные (вследствие наличия в фотической зоне постоянного светового градиента) для фитопланктонных организмов. Для некоторых бактериальных процессов водную среду также мож5 но считать, по крайней мере, несущественно структурированной. Не случайно всем известные более или менее крупномасштабные про5 цессы микроорганизмов (например, осуществляемые тионовыми бак5 териями) оказываются связанными именно с водной средой.
Все сказанное убеждает в том, что масштаб процесса пропорцио5 нален однородности среды (безразлично – отысканной организмом
вприроде или созданной человеком в лаборатории), соответствую5 щей по своим свойствам экологической нише определенного вида. Это соответствие требованиям вида, предъявляемым к окружающей среде, препятствует прекращению связанных с его жизнью процес5 сов. Как мы видели, в водной среде сравнительно большие участки биотопа могут быть рассмотрены как однородные по ряду свойств,
вкоторые «укладываются» ниши некоторых простейших организ5 мов. Другое дело – почва. Обилие в ней разнообразных градиентов обеспечивает ее высокую структурированность. Локусы в почве, со5 ответствующие точкам пространства ниши отдельных видов, как пра5
138

вило, весьма малы и тем самым обеспечивают поддержание вида в пределах микрооднородности, т.е. в виде пространственно не слиш5 ком протяженных популяций. Поэтому подавляющее число процес5 сов в почве является микропроцессами, локализованными микро5 однородностью ее структуры. Вполне естественно, что малые раз5 меры бактериальных ниш способствуют обнаружению неоднород5 ности биотопа и, следовательно, измельчению масштаба процессов, осуществляемых в границах локусов.
Таким образом, в пределах локуса масштаб процесса является функцией поддержания однородности изначальных условий, в ко5 торых процесс может начаться. Однородность обнаруживает свой5 ства протяженности в пространстве и времени. Однако, в последнем случае мы фактическим имеем дело лишь с временной аналогией размеров однородности среды.
Обсудим, как, собственно, велико разнообразие локусов, пригод5 ных для эксплуатации популяциями различных микроорганизмов?
Можно предположить, что по мере дробления каждого однород5 ного, с точки зрения макроструктуры, биотопа (например, луга) число локусов будет катастрофически расти (рис. 1, А). Альтерна5 тивное предположение допускает, что сходные локусы возникают в биотопах с разной макроструктурой. Поэтому с повышением макро5 структурного разнообразия биотопа микроструктурное разнообра5 зие не возрастает пропорционально, а может оставаться практически прежним. Вследствие этого при дроблении различных по макро5 структуре биотопов число различающихся по микроструктуре локу5 сов будет не так велико, как могло бы показаться на первый взгляд (рис. 1, Б). Поэтому количество видов, соответствующих разнооб5 разию локусов, не может быть безгранично большим.
В справедливости последнего предположения убеждает тот факт, что один и тот же микроорганизм встречается в биотопах, суще5 ственно различающихся по макроструктуре. Так, фотосинтезирую5 щие серобактерии даже одного вида можно обнаружить в лужах с дождевой водой, в водной толще озера и в придонном илу. Другим примером могут служить виды микроорганизмов, распространен5 ных повсеместно, – сенная и кишечная палочки или синезеленая водоросль Mycrocystis aeruginosa в сильно эвтрофированных и про5 гретых водоемах.
Поскольку выделение локуса связано с отысканием соответствия условий внешней среды требованиям вида, то сравнительная про5 стота экониш микроорганизмов, отмеченная выше, предопределяет и относительную простоту выделяемых локусов, т.е. некоторую их элементарность. Но всякое разнообразие элементарного ограничено больше, чем разнообразие сложного. Так, соединение примерно ста элементов таблицы Менделеева дает непредсказуемое множество раз5
139

личного рода соединений. Поэтому ограниченность разнообразия микроструктурированных локусов есть, в конечном счете, естествен5 ное следствие относительной простоты экологических ниш простей5 ших организмов.
До сих пор мы анализировали статические ситуации, решая прин5 ципиальные стороны соответствий, так что даже изменения во вре5 мени рассматривались как пространственные аналоги поддержания однородности. Введем теперь в рассмотрение динамический момент, чтобы обсудить некоторые возможные следствия соразмерности из5 менений внешней среды с генетической изменчивостью признаков вида, определяющих структуру его экологической ниши. Для упро5 щения последующего анализа примем две биологически правдопо5 добные предпосылки, не противоречащие концепции проб и ошибок.
1.Вероятность возникновения генетических проб за один и тот же отрезок времени связана обратной зависимостью с временем жизни одной генерации. При едином плане генетической организации от бактерий до человека время жизни организма микроскопических размеров и высокоорганизованной макроособи различаются на 6–7 порядков (у бактерий и водорослей минуты и часы, у птиц и млеко5 питающих десятки и даже сотни лет). Поэтому при едином времени геологического отсчета относительно возраста Земли микроскопи5 ческие формы жизни имеют вероятностные преимущества перед всеми другими, более сложными организмами в появлении новых признаков, даже при допущении прямой пропорциональной зависи5 мости частоты возникновения мутаций от времени. На самом деле вероятность возникновения полезного для выживания вида призна5 ка (свойства) должна быть несколько большей у видов с относи5 тельно простыми и небольшими нишами.
2.Естественный отбор более эффективен в стабильных условиях внешней среды, чем в период ее изменения. Действительно, прово5 цируемые изменением среды изменения генетического материала в отсутствие стабильных условий находятся в состоянии непрерывно5 го изменения.
Возвратимся, однако, к обсуждаемому вопросу и проанализиру5 ем с учетом принятых предпосылок возможные последствия раз5
личного темпа изменения среды обитания (Vс) и темпа изменения структуры экологических ниш (Vн).
Рассмотрим первый вариант, при котором скорость изменения
ниш обгоняет скорость изменения среды (Vн>>Vс), вследствие чего по отношению к ряду поколений окружающую среду можно рассмат5 ривать как стабильную. В этом случае повышенная вероятность по5 явления новых признаков и высокая эффективность их отбора в период относительной стабилизация внешних условий приводит в итоге к максимально достижимому соответствию ниш условиям локусов
140