

Изменения в природных биологических системах - Федоров В.Д
..pdfПринципиально новым по характеру явился другой показатель разнообразия  или
или  , предложенный Маргалефом [4]. Этот по5
, предложенный Маргалефом [4]. Этот по5
казатель учитывает массовость каждого вида анализируемого сооб5 щества. Расчет Е ведется по формуле:
(8)
где ni – численность i5того вида в анализируемой пробе, так что
Для достаточно больших значений ni и N при расчете факториалов удобно пользоваться формулой Стирлинга:
(9)
При отыскании lgN при N>100 эта формула может быть значи5 тельно упрощена:
(10)
Величина индекса Е зависит от объема исследуемой пробы, и поэ5 тому для сравнения проб различного объема удобно использовать
отношение  (или
(или 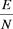 ), где Eмакс=lgN!. Отношение
), где Eмакс=lgN!. Отношение  меняется
меняется
от 0 в том случае, когда анализируемая совокупность организмов принадлежит к одному виду, до 1, когда все особи пробы принадле5
жат к разным видам. Отношение  в тех же условиях изменяется
в тех же условиях изменяется
от 0 до lgw. Последнее отношение при достаточно больших N (от 100 и выше) оказывается эквивалентным показателю биотического разнообразия, вычисленному по энтропийной функции Шеннона:
(11)
где  есть вероятность найти особь i5того вида среди сово5
есть вероятность найти особь i5того вида среди сово5
купности из N экземпляров. Этот переход от функции , записан5 ный как
(12)
легко понять, если иметь в виду, что величина Е (8) представля5 ет собой статистическую энтропию системы из N частиц, относя5 щихся к w сортам. (Ком. 3)
101

Показатель разнообразия D также меняется от 0 при w=1 до lg w при w=N. В наших пробах, собранных в течение сезона, макси5 мальное число учитываемых видов в 1 л воды равнялось 16, и по5 этому можно было ожидать изменения D по сезону в интервале от 0 до 1,2. Фактически, как видно из рисунка 2, величины D, сглажен5 ные по методу взвешенной скользящей средней, колеблются от 0,35 до 0,85 при средней квадратичной ошибке s2{D} = 0,0554. Легко видеть, что размах колебаний с 95%5ной вероятностью превосходит величину случайных отклонений при определении D.
Таким образом, показатель D, имеющий вполне определенное экологическое значение в смысле информационной энтропии Шен5 нона, оказывается достаточно чувствительным к структурным изме5 нениям однотипного сообщества во времени. Напомним, что при расчете a не учитываются различия в численности отдельных видов внутри сообщества, в то время как D является аддитивной величи5 ной, отдельные слагаемые которой определяются именно относи5 тельной численностью входящих в сообщество видов. В данном случае мы имеет два показателя, которые обнаруживают смысловые различия и формально пригодны для характеристики разнообразия сообщества. Так, если a выражает чисто формальную сторону связи между числом видов и общей численностью, вследствие чего биоло5 гическая интерпретация индекса Фишера представляется затрудни5 тельной, то показатель D имеет вполне реальный смысл, отражая меру неопределенности рассматриваемой системы. (Ком. 4)
Некоторой модификацией показателя биотического разнообразия
является отношение которое, меняясь от 0 до 1, заключает
в себе также информацию о близости системы к максимально воз5 можному разнообразию. Это отношение может иметь самостоятель5
ное значение, поскольку, в отличие от D, при расчете учитыва5
ется число видов, содержащихся в единице биотопа. В данном слу5 чае в скрытой форме показатель  зависит от объема (или площа5
зависит от объема (или площа5
ди) биотопа, выбранного в качестве единицы измерения. Действи5 тельно, при определении Dмакс следует пользоваться формулой (1) для определения числа видов w по суммарной численности N, полу5 чаемой пересчетом с пробы, в выбранной единице пространства био5 топа (1 см, 1 л, 1 м и т.д.). Так, например, в пробе № 1 1004 клетки, учтенные под микроскопом, принадлежали к 9 видам, и распреде5 ление их численности по видам формально удовлетворяло закону распределения Фишера.
Значение  , найденное по рисунку 1, оказалось равным 1,38. При пересчете на 1 л суммарная численность составила 33530 кле5
, найденное по рисунку 1, оказалось равным 1,38. При пересчете на 1 л суммарная численность составила 33530 кле5
102

ток, из чего следует, что в 1 л будет:
видов.
При величине D = 0,550 (рассчитано по формуле 12) отношение  равно 0,480. Однако следует оговорить, что достоверность раз5
равно 0,480. Однако следует оговорить, что достоверность раз5
личия между фактическим и рассчитанным по закону Фишера рас5 пределением может быть мала из5за небольшого числа видов в пробе.
Для исследованных нами проб с указанными выше интервалами N и w после построения диаграмм парной корреляции оказалось, что между показателями  и D отсутствует существенная корреляция,
и D отсутствует существенная корреляция,
а между 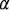 и обнаруживается отрицательная корреляция, осо5
и обнаруживается отрицательная корреляция, осо5
бенно отчетливая в области низких значений индекса Фишера (a<1,9). Отсутствие связи между  и D представляется вполне понятным, поскольку первый показатель характеризует разнообразие сообще5 ства по обобщенным признакам – суммарной численности и общему числу видов, тогда как второй отражает особенности относительного распределения видов по их численности. В общем случае нет основа5 ний постулировать обязательность проявления связи между указан5 ными особенностями структуры. Поэтому наблюдаемую отрицатель5
и D представляется вполне понятным, поскольку первый показатель характеризует разнообразие сообще5 ства по обобщенным признакам – суммарной численности и общему числу видов, тогда как второй отражает особенности относительного распределения видов по их численности. В общем случае нет основа5 ний постулировать обязательность проявления связи между указан5 ными особенностями структуры. Поэтому наблюдаемую отрицатель5
ную корреляцию между  и следует, по5видимому, рассматри5 вать как частный случай, а не как проявление общего правила. (Ком. 5)
и следует, по5видимому, рассматри5 вать как частный случай, а не как проявление общего правила. (Ком. 5)
Гораздо более интересным представляется отыскание связи между показателями разнообразия, отражающими структурные особеннос5 ти сообщества, и его важнейшей функциональной характеристикой – продуктивностью.
Маргалеф [5, 6, 7] исследовал наличие подобного рода связи между некоторыми структурными показателями (содержание хло5 рофилла а, пигментным отношением D430 : D665, биотическим разно5 образием D) и основными продукционными характеристиками – валовой (Рg, мкг, углевода на 1 л в час) и удельной (Pv, мкг углерода на 1 мкг биомассы) продуктивностью. Он нашел, что пигментное отношение и биотическое разнообразие отрицательно коррелируют (50,968 и 50,723 соответственно) с Рv. Слабая отрицательная корре5 ляция (50,107) была обнаружена между D и Pg. Это позволило Мар5 галефу заключить, что любое уравнение, дающее математическое описание продукционного процесса, должно включать некоторый член, количественно отражающий структуру сообщества.
Мы при сборе проб проводили также измерения продуктивности фитопланктона радиоуглеродным методом. Сезонные изменения Рg и Рv приведены на рис. 3.
103

Рис. 3. Сезонные изменения валовой (1) и удельной (2) продуктивности фитопланктонного сообщества
Сопоставление кривых Рg c D и  отчетливо демонстрирует
отчетливо демонстрирует
наличие обратной корреляции между этими показателями. Действи5 тельно, максимумы на одной из кривых почти точно совпадают по времени с максимумами на другой. Корреляционная диаграмма не
дает столь отчетливой картины связи между D и 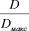 , с одной сто5
, с одной сто5
роны, и Рg, с другой, поскольку она не отражает изменений системы во времени. Однако, она может оказаться полезной для более точно5 го анализа полученных данных при условии, что фактор времени учитывается путем нумерации точек во временной последовательнос5 ти (рис.4). Легко видеть, что пробы летнего периода обнаруживают
максимум изменения  (от 0,81 до 0,24) и D (от 0,36 до 0,80)
(от 0,81 до 0,24) и D (от 0,36 до 0,80)
при сравнительно малом изменении продуктивности (от 4,0 до 5,0 мг углерода на 1 л в час). Подобного рода функциональная стабиль5 ность фитопланктонного сообщества в летний период соответствует, по5видимому, состоянию всей экосистемы, близкому к равновесию. В этом случае мы имеем право рассматривать структурные изменения однотипного сообщества как отражение данного механизма авторегу5 ляции (на основе обратной связи) который в течение этого периода обеспечивает функциональную стабильность сообщества за счет «ком5
104
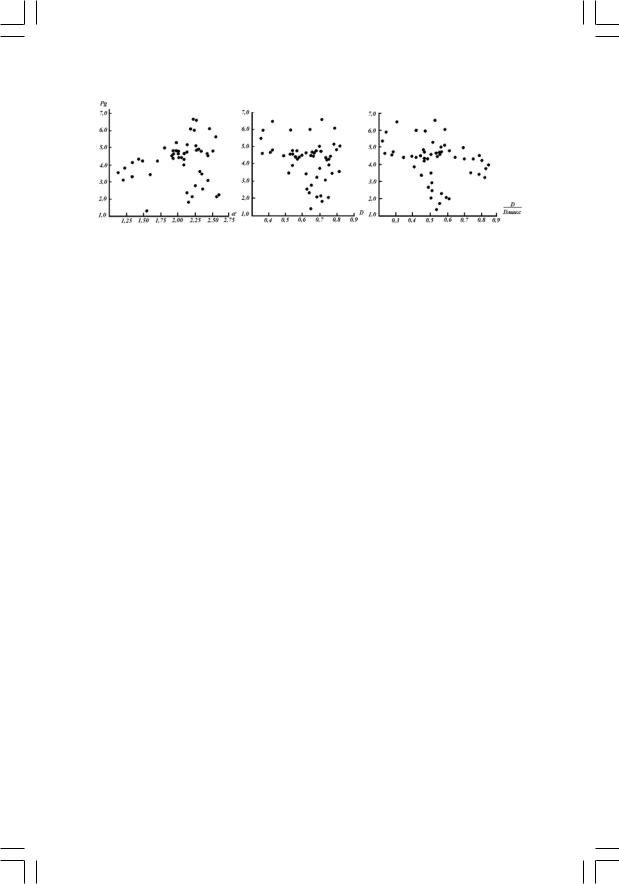
Рис. 4. Корреляция показателей разнообразия |
с |
валовой продуктивностью Рg |
|
пенсирующих» изменений численности составляющих его видов. Таким образом, в летний период между D,  и Рg теряет форму
и Рg теряет форму
простой зависимости (обратная корреляция). Иными словами, в период завершающей формации (климакса) структурные измене5 ния, измеряемые показателями информационной энтропии, направ5 лены на поддержание неизменности функциональных характеристик. В этом, собственно, и проявляется конкретный механизм сбаланси5 рованности системы в целом, т.е. механизм динамичного равнове5 сия между живой и неживой компонентами экосистемы.
Совершенно иной характер имеет корреляция Рg и  (рис. 4). В течение весеннего (за исключением проб № 1–4, отобранных подо льдом) и всего летнего периода обнаруживается положительная ли5 нейная корреляция (+0,666) между индексом Фишера и валовой про5 дуктивностью. Осенние точки и весенние подледные явно выпадают из общего пассива точек вследствие лимитирующего действия физи5 ческих факторов (световой обеспеченности и температуры) в эти пе5 риоды. (Ком. 6) Таким образом,
(рис. 4). В течение весеннего (за исключением проб № 1–4, отобранных подо льдом) и всего летнего периода обнаруживается положительная ли5 нейная корреляция (+0,666) между индексом Фишера и валовой про5 дуктивностью. Осенние точки и весенние подледные явно выпадают из общего пассива точек вследствие лимитирующего действия физи5 ческих факторов (световой обеспеченности и температуры) в эти пе5 риоды. (Ком. 6) Таким образом,  как показатель, не чувствительный к индивидуальным колебаниям численности отдельных видов сооб5 щества, по характеру сезонных изменений более близок к такому общему функциональному показателю, как валовая продуктивность.
как показатель, не чувствительный к индивидуальным колебаниям численности отдельных видов сооб5 щества, по характеру сезонных изменений более близок к такому общему функциональному показателю, как валовая продуктивность.
Сопоставление изменений удельной продуктивности с сезонны5 ми изменениями показателей разнообразия обнаруживает хорошую обратную зависимость между Рv и  (коэффициент криволинейной корреляции равен – 0,744. Положительная криволинейная зависи5
(коэффициент криволинейной корреляции равен – 0,744. Положительная криволинейная зависи5
мость между Рv и  выражена значительно слабее, тогда как меж5 ду Рv и D она не обнаруживается вовсе (рис. 5). Любопытно, что
выражена значительно слабее, тогда как меж5 ду Рv и D она не обнаруживается вовсе (рис. 5). Любопытно, что
минимальные значения |
и соответствующей им активности при5 |
5
105

Рис. 5. Корреляция показателей разнообразия |
с |
удельной (по биомассе) продуктивностью Рg
ходятся на пробы с абсолютным доминированием одного вида (Sce5 letonema costatum Cl.), численность которого близка к своему пре5 дельному значению. Наоборот, максимальные величины обоих пока5 зателей приурочены к пробам c самой низкой суммарной численностью фитопланктона (N 1000 кл/л) и почти равномерным распределени5
1000 кл/л) и почти равномерным распределени5
ем численности по видам (ω = 7–8). Неудивительно, что при столь
разреженной плотности заселения биотопа и отсутствии пищевой кон5 куренции между видами, соответствует первой модели Мак5Артура [2], фитопланктонное сообщество являет максимальную удельную активность на фоне относительно низкой валовой продуктивности. При сравнении диаграмм, изображенных на рисунках 4 и 5, хорошо заметно изменение характера положения на них точек, относящихся к пробам, отобранным в период состояния системы, близкого к рав5 новесному (пробы № 15–45). Вертикальное размещение этих точек
на диаграмме Рg – при логарифмическом преобразовании Рv
обращается в диагональное на программе lg Pv – |
. Следова5 |
тельно, структурные изменения состава в этот период связаны с изменением удельной активности единицы биомассы. В результате при значительных колебаниях биомассы сообщества величины ва5 ловой продуктивности стабилизируются.
В этих изменениях Рv и отображен, по5видимому, обсуждавший5 ся выше механизм авторегуляции Рg, т.е. механизм функциональ5 ной стабилизации сообщества.
Таким образом, продукционные характеристики в наших опытах оказывались тесно связанными с индексом разнообразия Фишера:
(13)
(14)
106

Для показателя  достоверная корреляция обнаруживается только после логарифмического преобразования Рv:
достоверная корреляция обнаруживается только после логарифмического преобразования Рv:
(15)
тогда как показатель биотического разнообразия D в целом по сезону не обнаруживает существенной корреляционной связи с про5 дуктивностью фитопланктонного сообщества.
Итак, индекс разнообразия Фишера, отражающий в большей мере внешнюю сторону сложности системы (учитывает общую численность и число видов и не учитывает характера распределения N по w5клас5 сам), обнаруживает более тесную связь с суммарными функциональ5 ными характеристиками системы (продуктивностью), чем показате5
ли биотического разнообразия – D и |
. Последние регистрируют |
лишь относительные колебания численности видов внутри сообще5 ства (т.е. внутреннюю сторону сложности системы), не учитывают изменений суммарной численности (N) и лишь слабо зависят от числа видов (w). Поэтому сравниваемые индексы разнообразия до5 полняют друг друга в описании системы на правах внутренних (D,
 ) и внешних (a) характеристик ее сложности. При этом каж5
) и внешних (a) характеристик ее сложности. При этом каж5
дый из них обнаруживает ту или иную связь с отдельными показа5 телями функциональной активности сообщества.
Л и т е р а т у р а :
Рудяков Ю.А. Применение индексов разнообразия в гидробиоло5 гических исследованиях // Труды Института океанологии. 1964. Т. 65.
Федоров В.Д. Организация сообщества фитопланктонных организмов и сезонные изменения его структуры // Ботанический журнал (в печати).
Fisher R.A., Corbet A.S., and Williams C.B. The relation between the number of species and the number of individuals in arandom sample on a animal population // Journ. Animal Ecol. 1943. Vol 12.
Margalef R. Temporal succession and spatial heterogeneity in phytoplank5 ton // Perspectives in Marine Biology. 1958. Univ. California Press.
Margalef R Modelos simplificados del ambiente marino para el estudio de la sucesion y distribucion del fitoplancton y del valor indicador de sus pigmentos // Inv. Pesq., 1963. T. 23, p. 11–52.
Margalef R. Correspondence between the classic types of lakes and the structural and dynamic properties of their populations // Verh. Int. Ver. Limnol., 1964. Vol. 15, p. 169–175.
107

Margalef R. Ecological correlation and the relationship between primary productivity and community structure // Men. Ist. Ital. Idrobiol., 1965. 18 Suppl., p. 355–364.
Биол. науки, №2, 1970.
1)Сейчас такими упражнениями никого не удивишь, но в те!то годы все эти вычисления делали вручную, точнее, с помощью арифмометра «Феликс»!
2)Не могу не отметить скрупулезность статистических оценок, полученных в те времена, когда никаких персональных компьютеров не было еще и в проекте, а вопрос, стоит ли заводить на Биологическом факультете ЭВМ, был предметом долгих дискуссий. А многие ли ны! нешние исследователи могут похвастаться тем, что в их работе фор! мулируются и проверяются все необходимые 0!гипотезы?
3)Теперь–то мы все знаем, что никакая это не энтропия и не надо было притягивать за уши теорию информации, но сколько за прошед! шие 30 лет было написано (и пишется до сих пор) всякой околонаучной ерунды в связи с пресловутой формулой Шеннона!Уивера! И, между прочим, W.Weaver имеет к этой формуле косвенное отношение, по! скольку был вместе с Клодом Шенноном редактором небольшой, но до сих пор часто цитируемой книги, изданной в 1949 г.: «The Mathematical Theory of Communication». В этой книге, по!видимому, и была впервые опубликована энтропийная формула. Самому Шеннону возможность ее использования в экологии, скорее всего и во сне не снилась. А виноват во всем Рамон Маргалеф, издавший 11 годами позже статью с громким названием «Теория информации в экологии», в которой и предложил формулу Шеннона для измерения разнообразия сообществ. Бьюсь об заклад, что никто из нынешних гидробиологов и экологов, использую! щих «энтропийный индекс разнообразия», упомянутых книг не читал.
4)Вряд ли автор будет теперь настаивать именно на такой интер! претации получившихся корреляций, но в то время она представлялась вполне правдоподобной, а главное — наводила на некоторые нетриви! альные предположения (см. ниже).
5)Впоследствии отношение D/Dmax получило широкую известность под названием индекса выравненности Пиелу. Недавно один очень ум! ный человек из Екатеринбурга, Е.Л.Воробейчик, обратил внимание на то, что этот, якобы энтропийный, индекс легко сводится к линейной комбинации коэффициента вариации и коэффициента асимметрии для распределения видов по обилию в любой случайной выборке. Иными сло! вами — никакой новой информации по сравнению с традиционными ста! тистическими методами индексы разнообразия не дают.
6)Вот в этом и состоит основная ценность данной работы: здесь впервые сформулирован главный принцип регуляции в экосистемах, под! робно обсуждаемый в более поздних работах автора. Нельзя не при! знать, что увлечение модными в то время индексами разнообразия
оказалось все!таки более полезным, чем этого можно было ожидать.
108

ПЕРВИЧНАЯ ПРОДУКЦИЯ КАК ФУНКЦИЯ СТРУКТУРЫ ФИТОПЛАНКТОННОГО СООБЩЕСТВА
Ранее нами была предложена функция I для характеристики фи5 зиологической активности экосистем
(1)
где ni – численность i5го вида в сообществе, nmi – рассчитанная предельная численность i5го вида в данном биотопе (2), w – общее число видов в сообществе.
Несмотря на то, что при расчет I используется один из показателей структуры сообщества (ni), сама функция I не должна рассматри5 ваться в качестве структурной характеристики системы, поскольку уже отношение ni /nmi отражает состояние i5ной популяции (ее фун5 кциональную гетерогенность) в однотипном сообществе. При этом постулируется, что изменение активности любой популяции, разви5 вающейся по логистической кривой, совпадает с изменение числа кле55
ток, отродившихся в единицу времени. Тогда выражение
отражает физиологическую активность каждой (i5й) популяции на определенном участке кривой роста, где ni/nmi – вероятность на5 хождения физиологически разноценных индивидуумов в каждый момент времени.
Наличие достоверных корреляций между продукционными харак5 теристиками сообщества (валовой продуктивностью и удельной по биомассе продуктивностью) и функцией I (1) показало пригодность предложенной функции для описания его функционального состоя5 ния. Найденные уравнения регрессии, связывающие эти показатели, сделали возможным расчет первичной продукции исключительно по данным обработки фитопланктонных проб. Однако, несмотря на то, что погрешность при расчете продукционных характеристик была статистически вполне удовлетворительной (коэффициент вариации 20–25%), казалось заманчивым «улучшить» уравнение, связываю5 щее I с продукционными характеристиками, введением каких5нибудь новых членов. Особенно перспективным представлялось введение до5 полнительных показателей, характеризующих структуру сообщества.
Рамон Маргалеф в 1965 г. отмечал, что в биологическом смысле продукция есть подлинная функция структуры сообщества; при со5 хранении последней его функция (в частности, продукция на едини5 цу биомассы) тоже остается постоянной. Любое изменение структуры сообщества в пространстве или во времени приводит к изменению в темпе продукции (4).
109

Мысль о тесной неразрывной связи структуры и функции фито5 планктонного сообщества безусловно верна и даже полезна, в осо5 бенности для исследователей, полагающих, что эта связь начинает проявляться где5то на уровне осетровых рыб и других более высо5 коорганизованных гидробионтов. Однако сам характер проявления этой связи, по5видимому, может быть более сложным, прежде всего для экосистем, находящихся в околоравновесном состоянии. В час5 тном случае для фитопланктона Белого моря было найдено, что в период биологического лета структурные изменения сообщества были направлены на поддержание его физиологической стабильности (5).
Тем не менее, отыскание дополнительных связей между продук5 ционными характеристиками и показателями структурного разно5 образия может послужить основанием для включения некоторых из них в уравнение множественной регрессии. При этом мы вправе
рассчитывать, что получаемые по такому уравнению величины бу5 дут ближе к значениям Р, найденным экспериментально.
бу5 дут ближе к значениям Р, найденным экспериментально.
Таблица 1
ПОКАЗАТЕЛИ КОРРЕЛЯЦИИ МЕЖДУ ПРОДУКЦИОННЫМИ И СТРУКТУРНЫМИ
ХАРАКТЕРИСТИКАМИ ФОТОПЛАНКТОННОГО СООБЩЕСТВА
r |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
r1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Pg |
|
|
|
|
|
|
|
|
– 0,155 |
– 0,213 |
0,547 |
|
0,109 |
|
|
0,321 |
|
– 0,287 |
||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Pv |
0,574 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– 0,708 |
– 0,707 |
0,351 |
|
0,724 |
|
|
|||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
lgPv |
0,435 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– 0,884 |
– 0,660 |
0,459 |
|
0,717 |
|
|
|||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
0,616 |
|
0,807 |
|
|
0,902 |
|
|
|
|
|
|
|
|
|
0,478 |
|
|
0,268 |
|
– 0,653 |
|||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0,404 |
|
0,742 |
|
|
0,732 |
|
0,621 |
|
|
|
|
|
|
|
|
|
– 0,402 |
– 0,755 |
|||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
D |
0,418 |
|
0,551 |
|
|
0,644 |
|
0,741 |
|
0,632 |
|
|
|
|
|
|
|
|
|
|
0,832 |
|
|
|||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
D/Dmax |
0,462 |
|
0,825 |
|
|
0,804 |
|
0,735 |
|
0,758 |
|
|
0,846 |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
П р и м е ч а н и е : Клетки оставлены пустыми там, где отыскание корре5 ляции лишено смысла.
110