

40. Условные рефлексы с двусторонней временной связью.
Применялись раздражители, каждый из
которых вызывает специфический
объективно наблюдаемый безусловный
рефлекс. При сочетании таких
раздражителей вырабатываются условные
рефлексыс двусторонней временной
связью, причем каждый из пары
сочетаемых раздражителей становится
условным сигналом для рефлекса
партнерного раздражителя, т.е. вызывает
его эффекторное выражение.
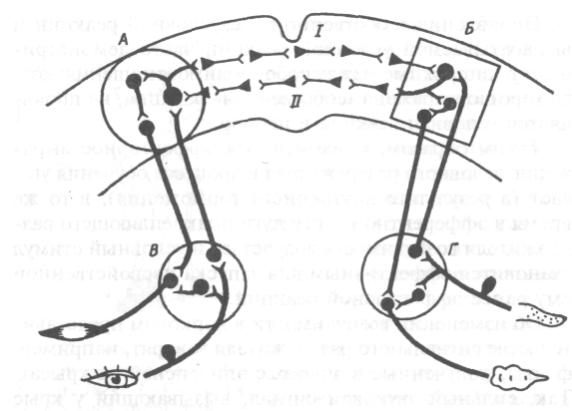
Схематическое изображение условного
рефлекса с двусторонней связью
А – кортикальный пункт мигательного рефлекса, Б – пищевого рефлекса, В и Г–
подкорковые центрымигательного и пищевого рефлексов
I – прямая временная связь, II – обратная
Возбуждение одного рефлекса тонически
меняет исходный функциональный фон
другого рефлекса. Под влиянием
внутреннего торможения с центра
безусловного рефлекса затормаживается
его среднее звено, в результате
редуцируется его специфическая
эфферентная реакция.
Образование условных рефлексов с прямыми и
обратными связями является общей закономерностью
«переключения» в высшей нервной деятельности.
Благодаря свойству условнорефлекгорного
«переключения» один и тот же раздражитель в
зависимости от обстановки опыта может приобретать
два или более различных сигнальных значения, каждое
из которых проявляется в адекватной ему ситуации.
Двусторонняя временная связь между кортикальными
«пунктами» условного и безусловного подкрепления
может служить субстратом для взаимодействия между
этими «пунктами», для их взаимного тонизирующего
влияния друг на друга, для циркуляции возбуждения
между ними и других взаимодействий.
41. Роль мозжечка в формировании условных рефлексов.
Мозжечок относится к многофункциональным
структурам мозга. среди его функций – сохранение
равновесия, поддержание позы, регуляция и
перераспределение мышечною тонуса, тонкая
координация произвольных движений.
В последние годы выявлена его способность
одновременно с корой формировать все виды
классических условных рефлексов. Благодаря связям
клеток Пуркинье мозжечка со всеми сенсорными
системами через мшистые, а затем через параллельные
волокна, а также с нижней оливой, откуда поступают
сигналы о всех совершаемых безусловных рефлексах,
клетки Пуркинье представляют уникальную основу
для конвергенции слоеного и безусловного сигналов.
Зачем в мозжечке параллельно
неокортексу формируются условные
рефлексы? Какую новую функцию по
сравнению с корой берет на себя
мозжечок? Предполагают, что мозжечок
контролирует точность выполнения
движений во времени, так как только
мозжечок обладает способностью в
любой момент времени заблокировать
любую двигательную реакцию или,
наоборот, дать ей возможность
реализоваться.
Причастность мозжечка к выработке классических
условных рефлексов доказывается опытами с
отключением холодом и разрушением его
структур. Временное охлаждение моторных
нейронов во время обучения блокирует выполнение
условного и безусловного рефлексов, но не нарушает
сам процесс обучения. С восстановлением функций
моторных нейронов можно обнаружить, что
процедура обучения в этих условиях завершилась
выработкой условного рефлекса, т.е. для процесса
обучения исполнение условной и безусловной
реакций необязательно. Если же во время выработки
условного рефлекса холодом отключить кору и ядра
мозжечка, обучение будет невозможным.
Частичным разрушением ядер моста, откуда берут
начало мшистые волокна, несущие афферентную
информацию к клеткам Пуркинье, можно вызвать
выпадение условных рефлексов на специфическую
модальность. Замена условного сигнала электрической
стимуляцией ядер моста приводит к более быстрому
формированию условного рефлекса. Разрушение
нижней оливы препятствует формированию
условных рефлексов в мозжечке.
Новые доказательства роли мозжечка в выработке
классического условного рефлекса получены на мышах-
мутантах, у которых через 2-4 недели после рождения
происходит полная дегенерация коры мозжечка. Такие
мыши способны лишь к частичному обучению.
Значительный прогресс в изучении
нейронных взаимодействий в мозжечке при
выработке классических условных рефлексов
был достигнут в работах Рихарда Томпсона,
доказавших, что условный мигательный
рефлекс формируется не только в коре, но
параллельно и в локальной зоне мозжечка.
В качестве условного сигнала он использовал
световой или звуковой раздражитель, а
безусловным рефлексом служило мигание
на обдувание воздухом роговицыкролика.
Основными клеточными элементами коры
мозжечка являются клетки Пуркинье. Их
дендриты восходят к поверхностным слоям, а
аксоны идут к нейронам ядер мозжечка. Кроме
клеток Пуркинье, в коре мозжечка имеются
зернистые и корзинчатые клетки. Афферентная
информация поступает в мозжечок к зернистым
клеткам от мшистых волокон (аксонов нейронов
моста). Клетки-зерна посылают параллельные
волокна к дендритам каждой клетки Пуркинье,
образуя на них множество синапсов. Параллельно
клетки-зерна образуют синапсы и на корзинчатых
клетках (интернейронах), оканчивающихся
тормозными синапсами на соме клетки Пуркинье.
Активация клеток Пуркинье создает торможение
у нейронов ядер мозжечка посредством ГАМК-
тормозного медиатора и как следствие -
торможение нейронов красного ядра,
управляющих двигательными рефлексами. На
дендриты клеток Пуркинье конвергируют не
только параллельные волокна зернистых клеток,
но и лазящие (или ползующие) волокна -
аксоны нейронов нижней оливы. Имеется около
15 млн клеток Пуркинье. Каждая из них имеет
возбуждающие синаптические контакты только с
одним лазящим волокном и много тысяч
синапсов – от параллельных волокон.
Таким образом, на дендритах каждой клетки
Пуркинье сходятся два потока влияний: от самых
разных афферентов (параллельные волокна) и от
нижней оливычерез одно лазящее волокно.
Сигнал, формирующийся в нижней оливе от
безусловного рефлекса, т.е. сигнал от подкрепления,
передается лазящему волокну, которое осуществляет
селективный выбор определенной клетки Пуркинье,
действуя по принципу: одно лазящее волокно - одна
клетка Пуркинье. Условный раздражитель представлен
на клетках Пуркинье возбужденным параллельным
волокном. Подобная конвергенция волокон имеет место
и на нейронах ядер мозжечка.
Обдувание воздухом роговицы возбуждает ядра
тройничного нерва и VI и VII пары ядер лицевых
мышц. Сигнал достигает мотонейронов, управляющих
движением мышц века, и вызывает мигательную
реакцию. От ядер тройничного нерва к ядрам лицевых
мышц идет дополнительный путь через ретикулярную
формацию, а также путь к мозжечку через нижнюю
оливу. Нейроны мозжечка имеют только один выход -
пучок аксонов клеток Пуркинье.
Установлено, что клетки Пуркинье характеризуются
тонической спонтанной активностью. Ее увеличение
означает усиление их тормозных влияний на нейроны
ядер мозжечка и красное ядро. Оно уменьшается со
снижением активности клеток Пуркинье. Тем самым
создаются условия для активации моторных центров и
появления двигательной реакции.
При выработке условного
рефлекса в мозжечке возникает
избирательное торможение
определенных клеток Пуркинье
по принципу «торможение
торможения», в результате чего
из-под тормозного контроля
высвобожается определенный
безусловный рефлекс.