

2.Етапи розвитку умовно-рефлекторної діяльності в філогенезі хребетних тварин.
Нервова система ланцетника, найпримітивнішої тварини з безчерепних, має дифузний характер, головний мозок фактично відсутній, а будова кожного з 62...64 сегментів нервової трубки приблизно однаково. Проте, не дивлячись на це, у них все ж таки вдається утворити тимчасові зв'язки (Б. Ф. Сєргєєв, 1962).
У ланцетника тимчасові зв'язки утворюються в тому випадку, якщо як умовні подразники використовуються підпорогові подразники (світлові, тактильні, електричні), а безумовним підкріпленням є ті ж подразники, що викликають чітку рухово-оборонну реакцію. Такі тимчасові зв'язки утворюються після 28...36 поєднань одномодальних подразників, зберігаються близько 10 г і згасають наступного дня.
Будова нервової системи круглоротих значно складніша. В їх головному мозку представлені всі основні відділи, існуючі у вищих тварин, хоча нервова система зберігає ще певні риси дифузної. У цих тварин можна утворити умовні рухові реакції на світло, вібрацію , тактильне і слабке електричне подразнення, причому для цього достатньо 3...11 поєднань. Вироблені умовні зв'язки відрізняються міцністю і часто зберігаються протягом декількох діб без додаткового підкріплення.
Проте умовнорефлекторні реакції безчерепних і круглоротих мають ряд особливостей. У них не виробляється умовна спеціалізована реакція на світло: заривання в пісок (для ланцетників) і присмоктування до скла (для міног) тільки в затемненій або освітленій частині акваріума. Крім того, єдиний вид внутрішнього гальмування, який можна у них виробити, — це згасаюче.
132
Таким чином, є всі підстави вважати, що у цих тварин виникає типовий тимчасовий зв'язок. Проте цей тимчасовий зв'язок має ряд особливостей.
По-перше, він утворюється переважно на підпорогові по силі подразники тих же модальностей, які викликають рухово-оборонну реакцію по типу безумовного рефлексу, хоча можливо утворення умовного рефлексу і при поєднанні різномодальних подразників, наприклад слабкого світла і сильного електричного струму або слабкого електричного струму і яскравого світла (90...120 люкс).
По-друге, утворення тимчасового зв'язку, навіть у разі використовування різномодальних подразників, не відбувається по типу замикання зв'язку між двома різними нервовими центрами. Тимчасовий зв'язок виявляється в підвищенні збудливості по відношенню до умовного подразника, тобто має певну схожість з сумаційним рефлексом, коли спостерігається підвищення збудливості до раніше індиферентного подразника. Коротше кажучи, на даному етапі еволюції утворення тимчасових зв'язків має дещо інший характер, ніж у більш високоорганізованих тварин.
На наступному етапі еволюції, у костистих риб, відбувається подальше ускладнення і диференціація нервової системи, виникає чітка локалізація центрального апарату основних видів чутливості — нюху, зору, органу бічної лінії і вестибулярного апарату, розвивається мозочок, тісно пов'язаний з центральною нервовою системою.
У костистих риб виробляються вже справжні умовні рефлекси на оптичні, акустичні, нюхові подразники, на постійне і змінне магнітне поле і інші прості і складні сигнали. Харчові і оборонні умовні рефлекси у риб згасають при непідкріпленні і «самовідновлюються» через деякий час. Отже, тимчасові зв'язки у них виникають при функціональному об'єднанні декількох нервових центрів, тобто є справжнім тимчасовим зв'язком. У риб також можливе утворення складних умовнорефлекторних реакцій з диференціюванням геометричних фігур і вибором місця підкріплення, а також різних видів внутрішнього гальмування.
Існує одна особливість тимчасових зв'язків у риб (а також у амфібій і рептилій), яка відрізняє їх від вищих тварин: неможливість синтезу компонентів складних подразників в єдине ціле, завдяки чому окремі компоненти комплексу не втрачають свого сигнального значення при ізольованому застосуванні.
Велика кількість досліджень присвячена вивченню різних поведінкових реакцій у амфібій. Найзвичайнішою поведінковою реакцією у безхвостих амфібій є звикання, обумовлене або згасанням орієнтувального рефлексу, або пов'язане з «негативним навчанням». За даними Б. Ф. Сєргеєва (1972), рухово-харчові умовні рефлекси у багатьох видів безхвостих амфібій виробляються за 3...12 поєднань і залишаються міцними за умови підтримання високого рівня харчової збудливості, а диференціювання
133
зорових харчових умовних подразників виробляється після 12...16 їх застосувань і зміцнюється після 15...18.
У жаб і ропух важко утворюються електрооборонні умовні рефлекси на включення загального освітлення, причому рефлекс виробляється тільки співпадаючий, при часі відставання підкріплення не більше 1 секунди. Це зв'язано з тим, що електричне подразнення не є достатньо адекватним стимулом, оскільки природжена оборонна реакція безхвостих амфібій в умовах деякого віддалення від видимого ворога, відсутності тактильних відчуттів схоплювання і достатнього простору для втечі виявляється в першу чергу в завмиранні. Не дивлячись на це, такий умовний рефлекс уникнення все ж таки виробляється у жаб, і після 75 поєднань світлового і електрошкірного подразників тварини правильно реагували в 66...93% випадків. Умовний рефлекс пасивного уникнення після його зміцнення зберігається протягом 10...35 днів (Б. Ф. Сєргеєв 1972).
У амфібій досить успішно відбувається «негативне» навчання. Так, наприклад, жаби і ропухи навчаються не їсти волохатих гусениць, а також черв'яків, змочених розчином хлористого кальцію або якщо при хапанні черв'яка через нього наноситься електричне подразнення. Крім того, ці тварини навчаються не їсти жалячих комах (бджіл і ос), і ця реакція зберігається до 2...3 тижнів (правда, є великі індивідуальні варіації). У безхвостих амфібій рухи схоплювання і заковтування з'єднані в єдиний комплекс, тому жаби важко навчаються не поїдати жалячих комах. Встановлено, що в цих випадках здатність до навчання залежить від здатності загальмовувати «автоматичну реакцію» кидка — «схоплювання — проковтування» — і «уклинити» в початок ланцюгової харчової реакції процес пізнавання, що вимагає якогось мінімального часу (Ю. Б. Мантейфель, 1977).
Таким чином, амфібії навчаються значно швидше, якщо умовним сигналом є екологічно адекватний подразник, а також на основі біологічно адекватної безумовнорефлекторної реакції.
Для оцінки рівня функціональної еволюції нервової системи звичайно використовують вивчення тимчасових зв'язків типу асоціацій по одночасності, тобто тимчасових зв'язків II типу, що виникають між індиферентними подразниками на базі орієнтовного рефлексу. В цьому випадку об'єднують між собою 20...150 разів два індиферентні подразники, а потім на один з них виробляють умовний рефлекс. Якщо другий (асоціативний) подразник набуває сигнального значення, аналогічного підкріплюваному, а контрольні подразники залишаються індиферентними, то це свідчить про встановлення тимчасового зв'язку II типу.
Порівняльно-фізіологічне дослідження різних груп хребетних тварин показало, що у ланцетників, міног, риб і амфібій тимчасові зв'язки II типу не утворюються. У рептилій вперше серед хребетних тварин з'являється здатність до утворення тимчасових зв'язків II типу, проте ці зв'язки виникають у них тільки при поєднанні одномодальних подразників. Що ж
134
стосується звичайних умовно-безумовних реакцій, то у рептилій можуть бути утворений різні екстероцептивні і інструментальні умовні рефлекси на базі харчової і оборонної реакцій.
На наступному етапі еволюції, у птахів, наявний високий рівень розвитку умовнорефлекторної діяльності. У них виробляються умовні рефлекси на предметні і слідові, одиночні і комплексні умовні подразники, імітаційні і умовні рефлекси II порядку, а також всі види внутрішнього гальмування. По багатьох показниках умовнорефлекторної діяльності птахи не відрізняються від ссавців, а деякі з них, наприклад воронові, по рівню розвитку елементарної розсудливої діяльності (екстраполяційний рефлекс) стоять вище за гризунів (кроликів, щурів) і знаходяться на одному рівні з собаками і кішками (Л. В. Крушинський, 1977).
Істинна здатність до утворення асоціацій між різномодальними подразниками вперше виникає у птахів. Такі тимчасові зв'язки II типу легко утворюються при 30...50 поєднаннях подразників, але при подальшому збільшенні кількості поєднань (до 75 ... 100) відбувається згасання тимчасовому зв'язку (Б. Ф. Сєргеєв, 1961).
Що стосується умовнорефлекторної діяльності ссавців, то вона вивчена вельми нерівномірно. Представники окремих рядів цього класу стали звичайним об'єктом лабораторного дослідження вищої нервової діяльності (собака, кішка, щур, мавпа і ін.). Значно менше досліджені тварини, що відносяться до інших рядів, і майже зовсім не досліджувалися підкласи клоачних і сумчастих.
У ссавців здатність до утворення асоціацій набуває подальшого розвитку, що пов'язано з прогресивним розвитком у них кортикоталамічної інтеграційної системи. У ссавців (за винятком гризунів) можливе утворення асоціацій по одночасності (умовних зв'язків II типу) збудливого і гальмівного характеру. У птахів і ссавців комплексні умовні рефлекси стають особливими синтетичними подразниками, компоненти яких з'єднані між собою системою тимчасових зв'язків. Синтез складних подразників, що складаються з
компонентів різних модальностей, і утворення асоціацій зв'язані у вищих ссавців з виникненням асоціативних областей кори (Б. Ф. Сергєєв, 1963). Так, в тім'яній асоціативній зоні кішок знайдена особлива група нейронів, що реагують тільки на поєднання двох або декількох подразників і не реагують на ізольовано вживані стимули.
Такі нейрони називаються «детекторами комплексу» і кількість їх складає близько 10% загального числа нейронів цієї ділянки мозку (В. А. Ізместьєв, 1975).
Роботами багатьох дослідників (М. М. Ладигіна-Котс, Р. Йеркс, В. Келлер, Е. Р. Вацуро, Л. А. Фірсов, А. И. Счастний і ін.), виконаними за допомогою методу умовних рефлексів і різних психологічних методик, було доказано, що мавпам властиві високорозвинута вища нервова діяльність, утворення різних позитивних і гальмівних умовних рефлексів і їх систем.
135
О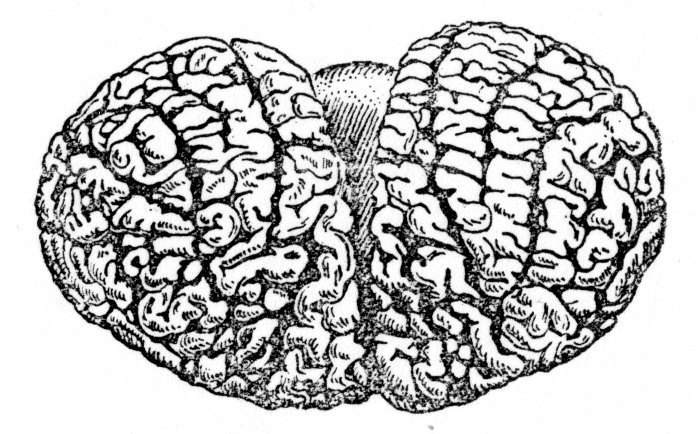 станнім
часом великий інтерес викликає поведінка
дельфінів, вивченню яких присвячено
багато досліджень. У дельфінів дуже
великий розмір півкуль головного мозку
і виражена складчастість кори (мал. 1).
станнім
часом великий інтерес викликає поведінка
дельфінів, вивченню яких присвячено
багато досліджень. У дельфінів дуже
великий розмір півкуль головного мозку
і виражена складчастість кори (мал. 1).
Мал.1. Фронтальна поверхня півкуль мозку дельфіна-афаліни.
У дельфіна-афаліни при масі тіла 70 кг маса головного мозку складає 1150 г, а маса великих півкуль рівна 81 % маси всього мозку (у людини відповідно 1400 г і 87%). Коефіцієнт теленцефалізації (відношення маси переднього мозку до маси всього мозку, виражене у відсотках) досягає у людини 87%, у дельфіна 75,6% і у антропоїдів 80%.
Відносна величина неокортексу складає: у дельфінів—97,8% поверхні всієї кори, у людини—95,5%, у шимпанзе— 93,3% і у собаки —84,5%. Проте, хоча у дельфінів нова кора займає до 98% всієї поверхні півкуль, лобова область — тільки 7,6...9,4% всієї кори. В лобових ділянках кори IV шар погано виражений, цитоархітектонічно ця зона поділяється тільки на чотири поля і нічим не відрізняється від лобової області хижих. Встановлено, що у дельфінів деякі параметри звукових і ультразвукових сигналів аналізуються на підкірковому рівні, а в кірковому відділі звукового аналізатора відбувається їх подальша переробка (М. Ф. Нікитенко, 1969).
Кора великих півкуль головного мозку дельфінів має шестишарову структуру і містить до 30 млрд. нейронів. Площа кори, прихована всередині звивин, у дельфінів складає 75...85% всієї поверхні кори (у людини 64...67%). Проте у дельфінів немає характерної для людини «колоної» організації кори, коли різного типу нейрони II—VI шарів розташовуються один під одним у вигляді вертикальних колонок, які можуть працювати самостійно або в комплексі з сусідніми.
Однією з помітних відмітних ознак кори мозку китоподібних є її рідкоклітинність і невелика товщина. Розділення кори на цитоархітектонічні шари виражено слабше, ніж у хижих або приматів, найбільш добре виділяється V шар з розміщеними мозаїчно і почергово ділянками згрупованих крупних пірамідних клітин і незвичайно широкий I шар.
Основну частину нейронів кори мозку дельфінів складають пірамідні клітини перехідного, проміжного типу, з нечітко вираженою формою. Орієнтація апікальных дендритів (особливо крупних пірамідних клітин V шару) нечітка: дендрити направлені не до поверхні кори, а під кутом або убік від білої речовини («перекинуті піраміди»). Дендритні шипики дуже великі і нечисленні. Клітини Беца, характерні для моторної зони кори у хижих і приматів, у китоподібних відсутні.
Дельфіни є аносматиками (у них відсутні нюхові цибулини і редукований нюховий нерв), але у цих тварин добре розвинутий нюховий мозок, що
136
складається з нюхових часток (палеокортекс) і відносно редукованого гіпокампу (архіпалеокортекс), а також мигдалевидного комплексу ядер. Отже, «нюховий мозок» (риненцефалон) у дельфінів виконує функції, не пов'язані з нюхом, і найімовірніше весь цей комплекс структур об'єднує передній мозок і ретикулярну формацію і забезпечує мотиваційно-емоційні реакції (У. Наута, 1968).
Навіщо дельфіну такий величезний мозок? Мабуть, провідну роль в розвитку такого мозку зіграла ехолокація. Крім того, сильному розвитку мозку у дельфінів сприяли такі чинники як велика швидкість плавання і швидка зміна умов при пірнанні; домінування звукового аналізатора, що сприймає акустичні коливання в діапазоні від десятків герц до 170 кГц; робота сонара (локатора) на високих частотах, яка дозволяє передавати відносно великий об'єм інформації за невеликий час; стадний і сімейний спосіб життя і сумісне полювання за рибою; тривале сумісне перебування дитинчат і батьків, що збагатило індивідуальним досвідом молодих особин (А. Р. Томілін, 1977).
Дослідження вищої нервової діяльності дельфінів-афалін методом умовних рефлексів показали значну інертність і неврівноваженість нервових процесів при хорошій їх концентрації. Це обумовлює швидке утворення тимчасових зв'язків і їх достатньо високу міцність (Л. Р. Воронін, Л. Б. Козаровіцький, 1969). На думку ряду дослідників, по швидкості вироблення різних умовно-рефлекторних форм поведінки дельфіни істотно не відрізняються від звичайних лабораторних тварин, наприклад, щурів (Ф. Бич, Р. Пеппер, 1972).
У дельфінів немає бінокулярного, стереоскопічного зору, проте вони здатні сприймати об'ємні предмети завдяки скануванню і обумовленому цим частковому перекриттю зорових полів, а також завдяки здатності висувати очі з орбіт.
Таким чином, тимчасові зв'язки значно розвинулися у філогенезі хребетних. Павлівський принцип можливості перетворення будь-якого подразника зовнішнього середовища в умовний сигнал будь-якої безумовнорефлекторної реакції зустрічає у нижчих хребетних певні обмеження. У них умовні рефлекси утворюються переважно на одномодальні подразники різної сили і не відбувається справжнього замикання тимчасового зв'язку між декількома нервовими центрами.
Проте, починаючи з риб, вже легко виникають різноманітні тимчасові зв'язки будь-яких зовнішніх подразників з безумовними реакціями, причому міцність і швидкість їх утворення залежать від екологічної адекватності подразника (Д. А. Відлюдків, 1948) і ступеня розвитку систем аналізаторів (Е. Р. Вацуро, 1948). Нарешті, у вищих тварин будь-які подразники зовнішнього (і внутрішньої) середовища організму легко вступають в тимчасовий зв'язок між собою.
Виникнення і розвиток нових форм тимчасових зв'язків завжди пов'язано з прогресивним розвитком нових структур мозку, але це не означає, що старі відділи нервової системи повністю втрачають цю здатність. Ймовірно, старіші відділи центральної нервової системи зберігають здатність до
137
утворення тимчасових зв'язків, по своєму характеру нагадують тимчасові зв'язки більш низькоорганізованих тварин.
Залежно від ступеня розвитку нервової системи і від умов життя умовнорефлекторна діяльність тварин різних класів, рядів, родин і навіть окремих видів в межах однієї родини має свої специфічні особливості. Проте протягом багатьох років при виробленні умовних рефлексів у різних видів тварин дослідники одержували в більшості випадків дуже схожі результати, внаслідок чого був зроблений висновок про те, що основні закономірності умовнорефлекторної діяльності однакові для різних тварин.
Дійсно, слід чекати схожості функцій у споріднених видів тварин, структура мозку яких дуже близька. Проте якщо взяти до уваги, що умовнорефлекторна діяльність є домінуючою функцією мозку, то б було дуже дивно, якби при чітких відмінностях в структурній організації мозку, наприклад, у риб і птахів, їх умовнорефлекторна діяльність істотно не відрізнялася.
Ці відмінності, як вже вказувалося, виявляються в здатності до утворення тимчасових зв'язків II типу і при утворенні звичайних умовно-безумовних рефлексів з різною величиною підкріплення. Так, наприклад, у щурів після зменшення величини харчового підкріплення умовного подразника спостерігається помітний неспокій і якийсь час вони виконують задачу повільніше, ніж тварини, які одержують із самого початку невеликі порції їжі. Цей ефект пригнічення в дослідах на рибах отримати не вдається.
Крім того, у щурів і голубів спостерігається обернено пропорційна, а у риб і черепах — пряма залежність між величиною підкріплення і швидкістю згасання умовної харчодобувної реакції.
Відмінності в умовнорефлекторній діяльності різних хребетних тварин визначаються якісними особливостями замикаючого механізму, а також домінуючим відділом головного мозку. У риб вищим інтеграційним центром поведінкових реакцій є середній мозок (А. І. Карамян), у рептилій і птахів — структури проміжного мозку і гіперстріатум (М. Ф. Поливана, Л. С. Ритікова), а у ссавців умовнорефлекторна діяльність є переважно функцією кори великих півкуль головного мозку (Д. А. Відлюдків, Л. Р. Воронін).
Необхідно наголосити, що не всі системи аналізаторів організму мають однакове, рівноцінне значення в умовно-рефлекторній діяльності. Тому у тварин, залежно від того, який з аналізаторів грає провідну роль в їх пристосувальній діяльності, в природних умовах існування переважають умовні рефлекси певного рецептивного апарату, і для них найхарактернішими (екологічно адекватними) є лише певні сигнальні
подразники.
На ці екологічно адекватні подразники умовні рефлекси виробляються швидше, легше і є дуже міцними. Наприклад, у зайця руховий умовний рефлекс на «чмокання» (звуковий подразник, що імітує його «голос») утворився після трьох поєднань подразників і був настільки міцним, що зберігався протягом півтора років, причому навіть 300 застосувань цього
138
сигналу без підкріплення не привели до повного згасання виробленої умовної реакції (Д. А. Відлюдків). В той же час руховий умовний рефлекс на звук метронома формувався значно повільніше, був дуже нестійким і повне його згасання відбувалося після 10 непідкріплень умовного подразника.
У водоплавних птахів найефективнішими подразниками є сплеск, а у бобрів — шум води, імітуючий прорив побудованої ними дамби. В той же час на інші звукові подразники умовні рефлекси у цих тварин виробляються значно повільніше. Встановлено, що у курей на умовний екологічно значущий сигнал (крик тривоги) реагує більше нейронів гіпокампу, ніж на звичайний умовний звуковий подразник (тон). Навіть величина умовної нейронної реакції на крик тривоги більше величини умовної реакції на тон (Ж. Р. Александрова, Н. Н. Василевський, 1977).
Ці особливості утворення умовних рефлексів необхідно обов'язково брати до уваги при вирішенні питання про швидкість утворення умовних рефлексів у хребетних різного рівня філогенезу. Дослідження Л. Г. Вороніна показали, що умовні рефлекси у всіх хребетних тварин (починаючи від риб і закінчуючи приматами) утворюються практично з однаковою швидкістю. В цьому немає нічого дивного, оскільки кожна тварина однаково добре пристосована до умов свого існування і тому, не дивлячись на відмінності в ступені розвитку нервової системи, пристосувальні реакції виникають однаково швидко у тварин різного рівня філогенетичного розвитку.
Відмінності, і іноді дуже істотні, у вищій нервовій діяльності різних хребетних тварин виявляються перш за все при здійсненні складніших форм аналітико-синтетичної діяльності або в спеціальних умовах експерименту.
Експериментально встановлено, що у всіх хребетних тварин можна виробити умовні рефлекси на комплексні подразники, причому швидкість вироблення таких рефлексів приблизно однакова. Наприклад, рухово-харчовий умовний рефлекс на одночасний комплексний подразник (світло електролампочки + дзвінок) утворився у риб, кроликів і мавп після 3...20 поєднань. Проте синтез компонентів комплексного подразника в єдине ціле у всіх трьох випадках був неоднаковий, що легко виявилося при виробленні диференціювання на ізольоване застосування окремих компонентів комплексного подразника, які не підкріплювались. У мавп таке диференціювання утворилося дуже швидко і легко (після 7 застосувань світла і дзвінка без підкріплення). Наявність позитивної реакції на комплексний подразник і гальмівна реакція на ізольовано вживані компоненти свідчать про те, що мавпи сприймають комплексний подразник як єдине ціле, а не як суму окремих компонентів, і на цей комплекс реакція зберігається повною мірою після згасання сигнального значення окремих компонентів.
У кроликів таке диференціювання утворюється значно важче, а у риб навіть після великої кількості непідкріплень окремі компоненти комплексного подразника не втрачають свого сигнального значення при їх ізольованому застосуванні. Якщо ж зробити навпаки, тобто позитивні умовні рефлекси
139
виробити на окремі компоненти, а комплекс з цих компонентів віддиференціювати шляхом його непідкріплення, то в цьому випадку не тільки у риб, але і у кроликів значно утрудняється аналіз і синтез комплексного подразника, оскільки таке диференціювання не вдається виробити, не дивлячись на велику кількість спроб.
У різних рядів ссавців також існують помітні відмінності в організації пристосувальної поведінки. Так, умовнорефлекторна діяльність комахоїдних (їжаків) характеризується досить низьким і примітивним рівнем синтетичних процесів і великою інертністю процесів збудження і гальмування. Це виявляється при виробленні умовних рефлексів на одночасний комплекс подразників (наприклад, світло + звук). Умовний рефлекс виробляється за 30... 40 поєднань. Проте при спробі гальмування окремих компонентів комплексу руховий умовний рефлекс на комплекс зникає, у їжаків розвивається тривалий агресивний невротичний стан. Крім того, у них дуже слабо розвинуті орієнтовно-дослідницькі реакції, тому будь-яка зміна обстановки досліду викликає оборонні реакції аж до невротичного зриву і відмови від продовження досліду (І. В. Малюкова, 1977).
Відмінності в рівні аналітико-синтетичної діяльності хребетних тварин виявляються ще більше при утворенні ланцюгових умовних рефлексів. Наприклад, у риб, черепах, курей, голубів, галок, ворон, кроликів, собак і мавп виробляли харчодобувні умовні рефлекси на трикомпонентний ланцюг подразників (дзвінок + метроном + світло). Швидкість утворення умовного рефлексу на такий подразник у різних тварин помітно не відрізнялася. Проте вже в самому процесі утворення рефлексу легко виявляється одна істотна відмінність. Так, якщо у ссавців відразу встановлюється зв'язок між безумовним подразником і окремими ланками ланцюга, то у риб, черепах і птахів спочатку утворюється зв'язок тільки з останнім компонентом і лише пізніше (після 5... 15 поєднань) виникає зв'язок з другим, а потім і з першим компонентом ланцюга.
Після утворення і зміцнення умовного рефлексу на ланцюговий подразник також виявляється помітна відмінність у взаємостосунках між його компонентами. У ссавців окремі компоненти при ізольованому застосуванні перестають викликати умовнорефлекторну реакцію. Особливо швидко (після 40...60 застосувань ланцюгового подразника) втрачають своє сигнальне значення окремі ланки ланцюга у мавп. У риб і черепах навіть після 1000...1300 застосувань ланцюгового подразника його компоненти не втрачають свого сигнального значення при ізольованому застосуванні (Л. Р. Воронін). Отже, чим вище рівень філогенетичного розвитку тварин, тим краще у них розвинута аналітико-синтетична діяльність мозку.
Таким чином, ускладнення структури і функцій центральної нервової системи в процесі філогенезу приводить до поступового вдосконалення вищої нервової діяльності у зв'язку з ускладненням взаємостосунків високорозвинутих тваринних організмів із зовнішнім середовищем, що
140
знаходить своє віддзеркалення перш за все в розвитку складних форм аналітико-синтетичної діяльності мозку.