

2. Молекулярна теорія пам’яті.
Молекулярна теорія пам'яті. Основною функцією нервової системи є отримання, зберігання і переробка інформації, що поступає, з навколишнього середовища, інтеграція її з придбаною раніше інформацією і, якщо потрібно, відповідь на неї відповідною адаптивною поведінкою.
Для засвоєння інформації мозку, як і машинам для обробки даних, потрібен код. Пошук такого коду був перш за все направлений на розшифровку біоелектричних процесів, оскільки передбачалося, що саме вони є основними носіями інформації. Проте, на думку А. Фессара (1968), ніякий електричний код не може зумовити довгострокову пам'ять, оскільки фіксована інформація не знищується повним припиненням електричної активності у разі гіпотермії або електрошоку. Найімовірніше, електрична активність мозкових структур грає певну роль при короткостроковій пам'яті шляхом реверберації нервових імпульсів в замкнутих ланцюгах нейронів.
Специфічність інформації міститься, швидше за все, в нейрональних взаємозв'язках і їх організації, а не в частоті імпульсациії або в її чіткому тимчасовому розподілі. Тому найбільш ймовірно, що в рамках системи мозкових ланцюгів відбувається хімічне кодування інформації, що поступає. Перші експерименти по біохімії пам'яті були проведені Е. М. Крепсом (1957) і X. Хіденом (1959), які показали зміни змісту РНК в мозку тварин, що навчаються. Надалі виявилося, що, дійсно, між силою умовнорефлекторного збудження і концентрацією РНК в корі і підкіркових структурах існує чіткий зв'язок (В. К. Болондінський, В. Д. Федоров, 1972; М. Р. Казахашвілі, М. В. Балавадзе, 1972; Н. А. Тушмалова і ін., 1974; Т.М.Семенова, 1971; X. Метьє, 1972).
Введення РНКази внутрибрюшинно, інтрацеребрально або на поверхню мозку щурів і кроликів викликало зникнення раніше вироблених умовних рефлексів і утрудняло вироблення нових (В. І. Арав, В. 3. Косарєва, 1971; Л. Г. Воронін і ін., 1968; Р. А. Данілова, 1969; І. І. Казеннова, 1972, 1974; О. А. Крилов, Р. А. Данілова, В. З. Тонгур, 1965; А. Н. Черкашин, А. А. Азарашвілі, 1972). Тим самим було показано, що РНК грає якусь важливу роль в механізмах збереження тимчасового зв'язку. Проте деякі дослідники отримали протилежні результати (Р. X. Божко, 1968; До. Гросс, Ф. Кері, 1965).
Окрім РНК, важлива роль в передачі пам'ятного сліду приписується і ДНК. Так, Дж. Гейто (1969) вважає, що саме ДНК грає основну роль в процесі навчання, а РНК тільки переносить нову інформацію, яка кодується на рівні білка. Б. Ф. Ванюшин і ін. (1974) показали, що в процесі вироблення рухового умовного рефлексу на світловий подразник в корі великих півкуль і гіпокампі щурів відбувається специфічне метилювання ДНК, причому зміни в метилюванні ДНК залежали від ступеня зміцнення умовного рефлексу і були найбільшими в процесі його вироблення.
103
Нарешті, ряд дослідників вважає, що інформація записується і зберігається в нервовій системі у вигляді білкової структури, а РНК необхідна тільки для передачі специфічного інформаційного коду (В. Ейді, 1970; X. Хіден, П. Ланге, 1972; Я. Буреш і ін., 1971). Ці висновки грунтуються на дослідах із застосуванням блокаторів білкового синтезу і-РНК: рибонуклеази, пуромицину і циклогексиміду (інгібіторів трансляції), а також актиномицину Д (інгібітору транскрипції ДНК і РНК). Застосування цих речовин в певні періоди після початку навчання запобігає консолідації пам'ятного сліду, тобто перехід короткострокової пам'яті в довгострокову (Б. Агранов, П. Клінгер, 1964; З. Барондес, Р. Коен, 1966, 1967; Дж. Флекснер і ін., 1963, 1967, 1971). Досліди з вивченням кількісних змін РНК і білка в процесі навчання і використовування інгібіторів білкового синтезу показали, що утримання інформації пов'язано із збільшенням синтезу РНК і білка. Крім того, для консолідації пам'ятного сліду необхідний нормальний рівень нейропередавачів, зокрема ацетилхоліну (Дж. Дойч, 1971) і катехоламінів (Р. Дісмукс, А. Рейк, 1972).
Важливим етапом у вивченні хімічної природи пам'яті були досліди з «передачею» пам'ятного сліду від одних тварин іншим, коли ін'єкція мозкових екстрактів від тварин-донорів, навчених якій-небудь нескладній навичці, полегшувала навчання тій же самій задачі тварин-реципієнтів (мал. 1).
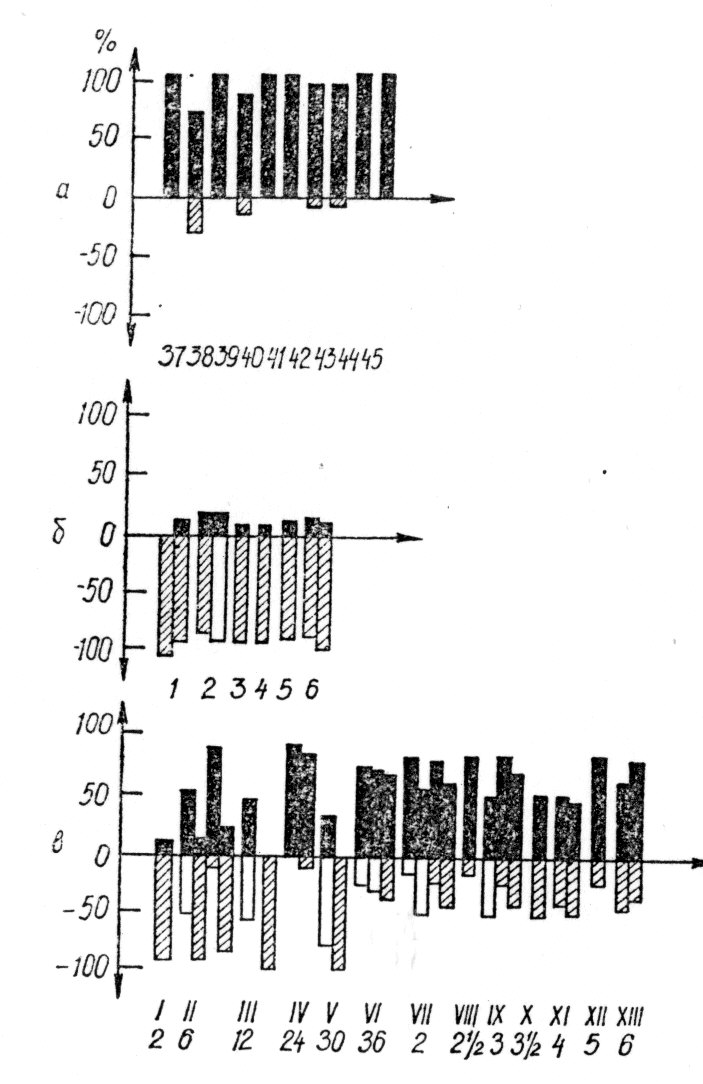
Мал. 1. Дія екстракту мозку риб-донорів на поведінку риб-реципієнтів (по Г. Домагк і Р. Зіппель, 1971):
а — вироблена реакція переважного руху до зеленого світла у риб-донорів; б — природжена реакція переважного руху до червоного світла у риб-реципієнтів до ін'єкції екстракту мозку риб-донорів; в —динаміка реакцій переваги до червоного і зеленого світла у риб-реципієнтів після ін'єкції екстракту мозку риб-донорів (від 2 г до 6 днів). Чорні стовпчики — реакція переваги до зеленої, заштриховані — до червоного і світлі — до білого світла.
Треба сказати, що такі | досліди, не дивлячись на їх уявну простоту, складні і отриманий результат залежить від багатьох змінних (вибору задачі, умови навчання, приготування екстрактів, дози ін'єкції, статі, віку і кількості тварин і т. п.). Крім того, виходить велика варіабельність результатів, оскільки ніякі два індивідууми не відповідають однаковою реакцією на один і той же подразник, що особливо яскраво виявляється при поведінкових пробах. Тому поведінка, індукована у реципієнтів, рідко буває точним повторенням навику, вивченого донором. Як правило, це тільки
104
бліда копія, чий зв'язок з оригіналом може бути точно встановлений тільки шляхом статистичної обробки даних і при порівнянні з відповідним контролем.
Тому часто позитивні результати, отримані в дослідах по «перенесенню пам'яті» за допомогою мозкових екстрактів, інтерпретувалися як наслідок загального полегшення консолідації пам'яті або неспецифічного стресу, обумовленого навчанням. Проте велика кількість (більше 200) експериментальих робіт свідчать про специфічність такого перенесення. Це дозволяє припустити наявність в мозковому матеріалі якоїсь досить специфічної інформації не тільки про винагороду, але і про спосіб її отримання.
Наші дані (Р. М. Чайченко, 1976) також свідчать про специфічність перенесення «пам'ятного сліду». Так, у однієї групи щурів був вироблений умовний харчодобувний рефлекс на дію звукового подразника, а в другій групі тварин здійснили гальмування орієнтовно-дослідницької реакції на цей же подразник, тобто тварини отримали стільки ж звукових подразників, як і щури першої групи, але без харчового підкріплення. Через 16 днів (80 застосувань подразників) з мозку обох груп щурів екстрагували РНК феноловим методом і вводили внутріочеревинно двом групам щурів-реципієнтів (з розрахунку 1:1). Третя група тварин була контрольною. Всі три групи щурів-реципієнтів ніколи не поміщалися в експериментальну камеру до введення їм РНК. Вибір щурів був випадковим; дослідження проводилося всліпу: експериментатор не знав, якій групі щурів і яка РНК була введена.
Перше випробування після ін'єкції РНК було проведено через 14 годин і ніякої різниці між трьома групами тварин в їх реакції на звуковий подразник знайти не вдалося. Але подальший дослід (через 38 г) виявив істотну різницю в поведінці щурів, що отримали екстракт мозку «навчених щурів» (I група). Тварини-реципієнти цієї групи активно здійснювали харчодобувну реакцію на дію умовного сигналу, тоді як щури, що отримали екстракт мозку донорів, у яких була загальмована орієнтовна реакція на цей сигнал (II група), і контрольні тварини (III група), яким вводили лише фізіологічний розчин, поводилися пасивно, ніяк не реагуючи на дію умовного подразника, або у них виникала пасивно-оборонна реакція.
В ході дослідження у щурів I і III груп був утворений стабільний умовний рефлекс, але у щурів I групи цей рефлекс утворився в 3 рази швидше, ніж у контрольних тварин III групи. Що стосується щурів II групи, що отримали РНК мозку щурів із загальмованим орієнтовним рефлексом на дію звукового подразника, то у 44% цих тварин умовний рефлекс не утворився навіть після 80 поєднань умовного і безумовного подразників (у контрольних тварин для цього знадобилося близько 20 поєднань) (мал. 2).
Після перерви в навчанні протягом 34 днів збереження виробленого умовного рефлексу склало: у щурів I группы— 97,5%, у щурів II групи (у
105
яких
цей рефлекс утворився)— 64,0% і у щурів
III
групи
(контрольних)-— 6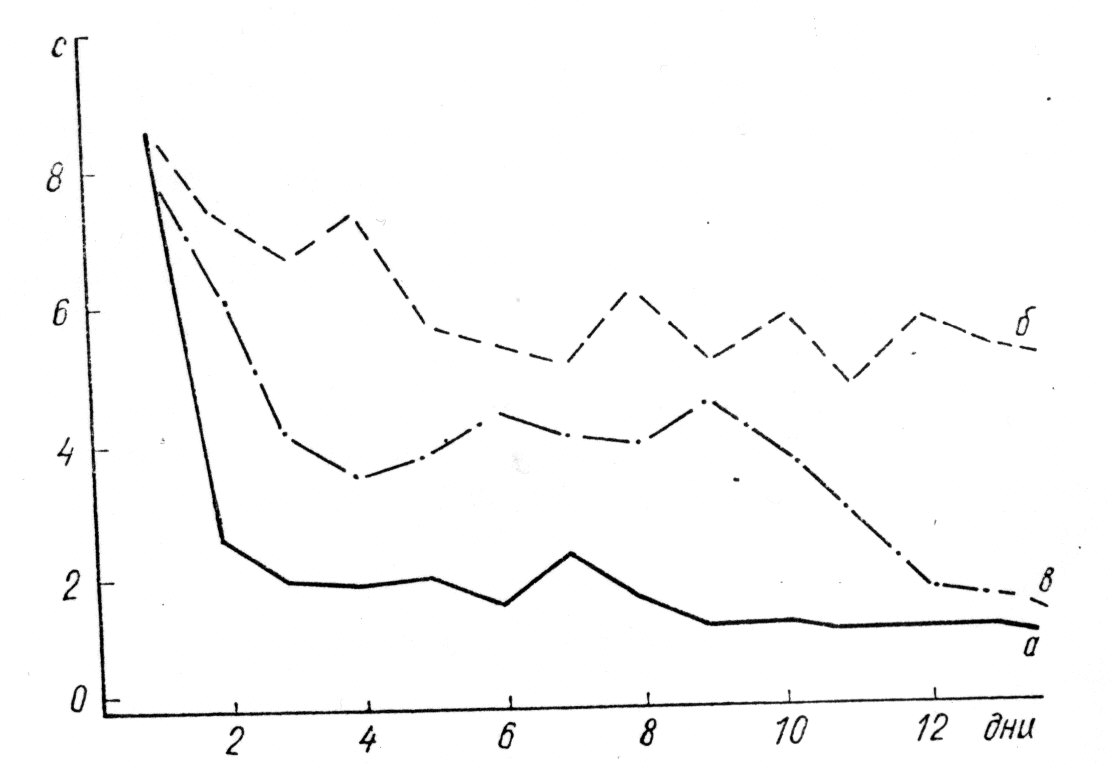 5,7%.
5,7%.
Мал. 2. Динаміка вироблення умовного харчодобувного рефлексу на звуковий подразник у щурів-реципієнтів, яким вводили РНК мозку щурів-донорів з виробленим рефлексом (а) і щурів-донорів, у яких була загальмована реакція на умовний звуковий подразник (б); в — контроль. По осі абсцис показані експериментальні дані, по осі ординат — величина латентного періоду умовної реакції (с).
Таким чином, експериментальні дані свідчать про специфічність перенесення пам'ятного сліду за допомогою мозкових екстрактів. РНК, ймовірно, служить лише одною, можливо дуже важливою ланкою, яка переносить інформацію до молекул білка, тобто довгострокова пам'ять обумовлена цілим комплексом структурних змін макромолекул.
Досліди Г. Унгара на щурах (1966—1968) при виробленні у них умовного рефлексу уникнення темноти показали, що активною речовиною є пептид, який можна витягнути з мозку методом холодної фенолової екстракції спільно з РНК. Цей єдиний комплекс (РНК плюс білок) розділяється діалізом при низькому рН (Р. Унгар, Е. Фьюрдінгштадт, 1971).
Отриманий ізольований пептид був названий скотофобіном (від грець. «боязнь темноти»), синтезований і перевірений з позитивним результатом в різних лабораторіях. Виявилося, що скотофобін дійсно викликає стресову реакцію у тварин, коли вони знаходяться в темноті (Д. Малини, 1974).
Подібним же чином в лабораторії Г. Унгара виробили у щурів звикання до звуку електричного дзвінка, а потім екстрагували і очистили активну речовину з їх мозку. Вона була названа амелитином (від греч. «байдужий»), синтезована і перевірена на свою ефективність. Виявилося, що дія синтетичного амелитину була дещо слабша, ніж природного продукту.
Спеціальні методи дослідження показали, що синтетичний скотофобін з'являється в мозку через 15 мхв після внутрішньоочеревинного введення, кількість його зростає протягом 2...3 г і потім поступово зменшується. Його вже не можна визначити через 48 г, хоча поведінковий ефект саме у цей момент досягає піку. А при виробленні у щурів навику уникнення темноти кількість природного скотофобіну зростає в перші 6 днів навчання, потім поступово знижується і через 15 днів знайти його в - мозку вже неможливо. Ймовірно, цей пептид зв'язується з яким-небудь синаптичним елементом і не може бути визначений існуючими методами (Р. Унгар, 1977).
106
Найбільша кількість скотофобіну і амелитину виявляється в корі (72,0 і 53,3% відповідно), менше — в мозочку (26,1 і 32,8%) і найменша кількість — в підкіркових утвореннях (1,9 і 13,9%).
До 1978 р. було виділено ще 3 пептиди, що обумовлюють специфічність колірної дискримінації і пристосовність до плавання із спеціальним поплавком у золотих рибок. Звичайно, важко допустити, що в мозку міститься достатня кількість пептидів, щоб забезпечити всю інформацію, накопичувану протягом життя і оцінювану в 1015 бітів (Р. фон Нейман, 1958), хоча якщо ці пептиди мають 15 амінокислотних залишків, як скотофобін, то можливо 2015 (тобто 3 х1019) пентадекапептидних ланцюжків, тобто значно більше необхідної кількості. Проте досліди показують, що після консолідації інформації вміст специфічного пептиду стає дуже низьким.
Отже, вираження пам'яті залежить від змін білків, які починаються і підтримуються якісними і кількісними змінами інформаційної РНК, що обумовлене процесом навчання. Втрата цієї специфічної РНК повинна привести до втрати необхідного (специфічного) пептиду з подальшою тимчасовою втратою пам'яті. Крім того, у присутності інгібіторів білкового синтезу концентрація специфічного білка може знизитися до рівня, дуже низького для вираження пам'яті, але ця амнезія повинна бути тимчасовою, якщо зберігається інформаційна РНК, щоб направити синтез білка після припинення дії інгібітору. Таким чином, РНК належить дуже важлива роль в процесі навчання і пам'яті.
Г. Унгар (1977) пропонує таке пояснення механізму консолідації пам'ятного сліду. Згідно гіпотезі Р. Сперрі (1963) про хемоспецифічність, нейрони володіють хімічними мітками, по яких вони взнають один одного і з'єднуються для утворення специфічних провідних шляхів, які формуються в ембріональному періоді і називаються протоланцюгами.
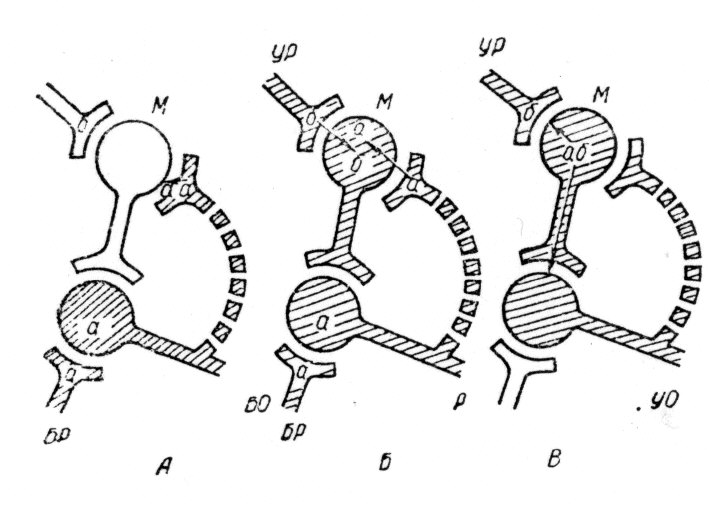
Мал. 3. Гіпотетичний механізм
формування металанцюгів (по Р. Унгару, 1977):
Л — безумовний рефлекс; Б — вироблення умовного рефлексу із залученням нейрона М, куда вводяться мітки «а» і «б»;
В — вироблений умовний рефлекс і утворення нового металанцюга з новим пептидом «аб», консолідуючим тимчасовий зв'язок.
БР — безумовний, УР — умовний подразник; УО — умовна відповідь;
М — спеціалізований інтернейрон («клітина пам'яті»). Заштриховані нейрони знаходяться в активному стані.
Найважливішою межею структурного коду є пластичність, тобто здатність себе постійно репрограмувати у міру надходження всякої нової інформації або ж формувати нові синаптичні зв'язки, не передбачені генетичним кодом.
107
Все це приводить в результаті до створення нових ланцюгів нейронів — металанцюгів, в яких і записується інформація.
Основні положення гіпотези Г. Унгара наступні (мал.3):
1. Активація провідних шляхів, залучених в навчання, веде до зростаючого синтезу їх генетичних міток.
2. Одночасні розряди в цих провідних шляхах викликають транссинаптичне перенесення міток «а» і «б» в спеціалізовані нейрони («клітини пам'яті»).
3. Ці клітини володіють ферментами (транспептидазами), здатними комбінувати «а» і «б» в новий пептид «аб». Так кодується нова інформація негенетичного походження.
4. Упровадження знов сформованого пептиду в активовані синаптичні мембрани приводить в результаті до створення металанцюга.
На думку Дж. Гейто (1976), найперспективнішим в даний час є підхід, направлений на виявлення якісної специфіки пов'язаних з навчанням макромолекул. Для цієї мети можуть бути використано 3 методи: метод гібридизації ДНК і РНК для виявлення специфічних РНК, метод електрофорезу на поліакриламідному гелі і експерименти по перенесенню навиків для виявлення специфічних білків.
Дослідження по відтворенню умовнорефлекторної діяльності при введенні екстрактів мозку від навчених тварин ненавченим достатньо переконливо свідчать про кодування інформації, про індивідуальний досвід на рівні макромолекул, хоча і не всі дослідники з цим згодні. Наприклад, М. М. Ліванов і З. І. Розанов категорично заперечують таку можливість, вважаючи, що пам'ятний слід зберігається виключно на основі електричної активності нейронних популяцій мозку.