

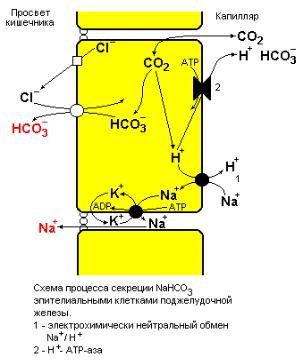
Современный способ лечения холеры основан на оральном введении раствора, содержащего 110 mM глюкозы, 99 mM Na+, 74 mM Cl¯ , 39 mM НСО3¯ и 4 mM К+. Данный способ разработан исходя из того факта, что совместный транспорт глюкозы и Na+ (см. лекцию 6) не зависит от сАМР и аденилат-циклазного пути.
Рис. 1.11 Модель эпителиальной секреции NaHCO3.
1 – электрически нейтральный обмен Na+/H+, 2 – Н+-АТР-аза.
Транспортная система, обеспечивающая всасывание глюкозы при заболевании холерой не страдает. Поэтому данный подход обеспечивает поглощение энтероцитами ионов Na+ в присутствии глюкозы и восстановление в организме потерь NaCl, связанных с болезнью. Основное преимущество предложенного метода терапии холеры состоит в простоте применения и низкой стоимости по сравнению с внутривенным введением жидкости (Carpenter C.C.J., Secretory diarrhea, Field M., Fordtran J.S., Schultz S.G., (eds), American Physiol. Society, MD: Bethesda, 1980, 67-83).
Секреция соляной кислоты обкладочными клетками желудка
Известно, что обкладочные клетки экзокринных желез желудка вырабатывают соляную кислоту, которая выбрасывается в полость желудка. При этом концентрация протонов (Н+) в полости желудка может достигать значения, равного 0,14М, что составляет величину рН желудочного сока, равную 0,8. Поскольку величина рН плазмы крови равна 7,4, обкладочные
клетки должны переносить протоны против градиента концентрации, разница которой равняется 10 6.6.
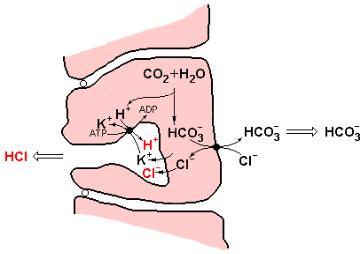
В процесс секреции соляной кислоты включается К+-зависимая Н+- АТРаза (К+,Н+-АТРаза). Этот фермент является уникальным для обкладочных клеток желудка и локализован только на апикальной стороне плазматической мембраны. К+,Н+-АТРаза связывает (сопрягает) процесс гидролиза АТР с обязательным электростатически нейтральным обменом К+ и Н+, продуцируя выброс протонов и поступление ионов К+ в клетку.
Рис. 1.12 Модель секреции соляной кислоты обкладочными клетками стенки желудка.
Встационарном состоянии HCl может вырабатываться в соответствии
сданным механизмом только в том случае, если апикальная часть мембраны проницаема для К+ и Cl¯, а базальная часть мембраны обеспечивает обмен
Cl¯ и НСО3¯. Обмен Cl¯ и НСО3¯ необходим для постоянного поступления в клетку ионов Cl¯ и для предотвращения защелачивания цитоплазмы. Таким
образом, в стационарных условиях, секреция HCl в полость желудка должна быть сопряжена с переносом НСО3¯ в плазму крови.
Лекция 2
Обмен белков. Общие положения
В процессах обмена веществ обмен белков занимает ведущее место, поскольку мономерные звенья пищевых белков – аминокислоты, в первую очередь, служат строительным материалом для любой клетки. Аминокислоты пищевых белков в равной степени необходимы как для продукции пищеварительных ферментов (многочисленные протеиназы желудочно-кишечного тракта, внутриклеточные протеиназы и пептидазы), принимающих участие в процессах переваривания пищи, так и для синтеза пептидных гормонов, осуществляющих тонкую регуляции функций разных систем организма. Пищевые белки необходимы для последующего синтеза протеинов плазмы крови, которые принимают участие в поддержании онкотического (осмотического) баланса, а также для синтеза белковпереносчиков малых молекул, в том числе и сигнальных молекул. Роль белков в функционировании иммунной системы также сложно переоценить. В целом белки осуществляют все метаболические процессы клетки и всего организма, выполняя уникальные каталитические функции.
Белки, как компоненты пищевых продуктов, выполняют также энергетическую функцию. Бóльшая часть аминокислот, так называемых глюкогенных аминокислот, превращается в процессе метаболизма в глюкозу. Другая часть аминокислот – кетогенных аминокислот – превращается в оксикислоты и жирные кислоты. Последние служат структурными элементами для синтеза триацилглицеролов, которые накапливаются в адипозной ткани. Однако роль и значение белков в процессах обмена веществ совершенно не определяется их энергетической ценностью. Энергия, получаемая при распаде белка, может быть без всякого ущерба для организма компенсирована энергией распада жиров и углеводов. Важно другое – организм человека и животных не может обходиться без регулярного поступления белков извне. Опыты на лабораторных животных показывают, что даже довольно длительное исключение жиров и углеводов из рациона не вызывает каких-либо тяжелых расстройств метаболизма и, соответственно, не влияет на состояние подопытных животных. Но кормление их в течение нескольких дней пищей, не содержащей белка, приводит к серьезным метаболическим сдвигам, а продолжительное безбелковое питание неизбежно заканчивается гибелью животного.
Таким образом, без белка, без составляющих его аминокислот не может быть обеспечено воспроизводство основных структурных элементов клеток, тканей и органов, а также образование ряда важнейших макромолекул, таких как ферменты, пептидные гормоны, иммуноглобулины, транспортные белки и многие другие.