

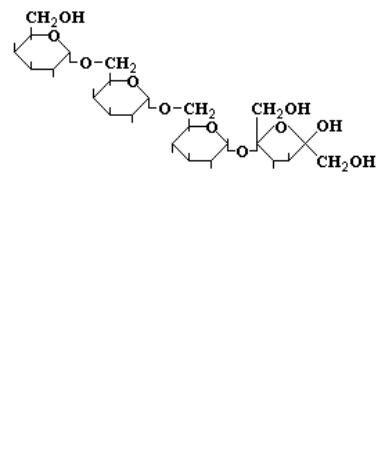
[α- Gal (1→6) α- Glc (1→2) β- Fru]
Почти так же широко, как сахароза и раффиноза в продуктах растительного происхождения распространен еще один олигосахарид – стахиоза, расщепление которого зависит от деятельности кишечной флоры:
[α- Gal (1→6) α- Gal (1→6) α- Glc (1→2) β- Fru ]
Молодые грибы часто содержат довольно большие количества необычного дисахарида (см. табл. 6.2) трегалозы [α-Glc (1→1) α-Glc], расщепление которого также обеспечивается только специфическим ферментом кишечных бактерий – трегалазой.
Всасывание моносахаридов
Впроцессе переваривания пищи мономерные звенья биополимеров (моносахариды, свободные аминокислоты) всасываются через эпителиальные клетки кишечника против градиента концентрации. Источником энергии, необходимой для такого «активного» транспорта,
служит не химическая энергия гидролиза АТР, а концентрационный градиент (или электрохимический потенциал) ионов Na+, создающийся на апикальной стороне плазматической мембраны. Примером такого
транспорта служит перенос глюкозы, который непосредственно зависит от электрохимического градиента ионов Na+ и только косвенно зависит от АТР.
Вусловиях in vivo глюкоза (и структурно родственные моносахариды) всасывается из полости кишечника в кровь через эпителий против градиента
концентрации. Этот направленный перенос является результатом сочетания нескольких отдельных мембранных процессов:
–АТР-зависимого переноса Na+ за пределы клетки, что служит причиной возникновения электрохимического натриевого потенциала;
–присутствия двух разных транспортных систем для глюкозы в апикальной и базальной частях плазматической мембраны энтероцитов;
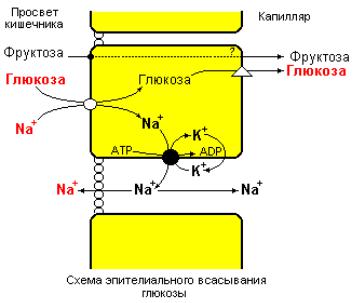
–сопряженного транспорта Na+ и глюкозы через апикальную сторону мембраны (рис. 6.3).
Рис. 6.3 Модель Na+-зависимого всасывания глюкозы и Na+-независимого всасывания фруктозы эпителиальными клетками тонкого кишечника
(Textbook of Biochemistry with clinical correlations, Devlin T.M., (ed.), WILEY-LISS, 1993).
Основная часть моносахаридов, образующихся в результате переваривания поли- и олигосахаридов, представлена, в первую очередь, D- глюкозой, затем D-галактозой и D-фруктозой. Всасывание этих и других, минорных, моносахаридов представляет собой процесс, зависящий от функционирования специфических переносчиков, которые характеризуются такими особенностями, как субстратная специфичность, стереоспецифичность, кинетика насыщения и отношение к специфическим ингибиторам.
В настоящее время известны, по крайней мере, два типа переносчиков моносахаридов, которые катализируют поступление этих углеводов из полости кишечника в энтероциты. Это, во-первых, переносчик, состоящий из тетрамеров с молекулярной массой около 75.000 дальтон/субъединицу, который обеспечивает совместный активный транспорт моносахаридов и

ионов Na+. Данный переносчик ответственный за транспорт D-глюкозы и D- галактозы специфически идентифицируется ингибитором флоризином. Вовторых, это Na+-независимый переносчик, обеспечивающий транспорт D- фруктозы по принципу облегченной диффузии.
Итак, апикальная сторона плазматической мембраны энтероцитов содержит транспортную систему, которая способствует тесно связанному переносу Na+ и D-глюкозы и структурно похожих углеводов в клетку. Данная транспортная система обеспечивают перенос глюкозы и Na+ в обоих направлениях в равной степени эффективно. Однако, вследствие более высокой концентрации ионов Na+ в полости кишечника и отрицательного потенциала в клетке (-60mV) совместный перенос Na+ и глюкозы происходит в направлении полость → клетка, даже не смотря на то, что концентрация глюкозы в клетке выше, чем в полости кишечника. Модельные эксперименты в условиях in vitro показывают, что разница в концентрации глюкозы в полости и в клетке может достигать двадцатикратной, если заблокировать отток глюкозы из клетки, например, специфическим ингибитором цитохалазином В.
Отсюда следует, что в ряде случаев именно поглощение Na+, описываемое данным механизмом, может быть наиболее важным для