

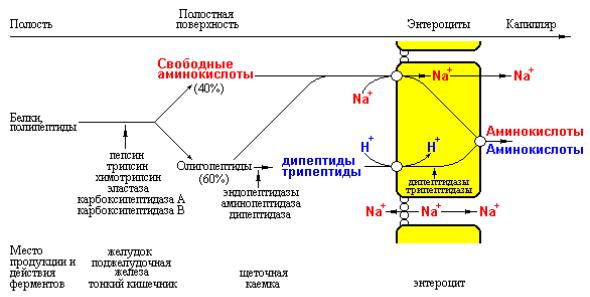
Рис. 3.20 Схема последовательных стадий расщепления белков до аминокислот и коротких пептидов в желудочно-кишечном тракте, а также Na+- зависимого транспорта свободных аминокислот и Н+-зависимого переноса ди- и трипептидов в энтероциты.
С другой стороны, всасывание ди- и трипептидов зависит от функционирования Н+-зависимой транспортной системы. Эти пептиды подвергаются расщеплению под действием олигопептидаз энтероцитов до свободных аминокислот, которые транспортируются в кровь с участием Na+- независимых систем базальной мембраны эпителиальных клеток кишечника.
Всасывание интактных белков
Эпителиальные клетки тонкого кишечника плода и новорожденных способны всасывать интактные белки. Поступление белков в клетки происходит посредством эндоцитоза. Этот процесс более правильно называть пиноцитозом из-за небольших размеров везикул поступающих внутрь клетки. Полагают, что пиноцитоз белков клетками тонкого кишечника чрезвычайно важен для осуществления переноса антител матери (γ-глобулинов) в организм плода и новорожденного. Пиноцитозное поступление белков не существенно для питания и, поэтому, после рождения эффективность процесса снижается. Однако продолжение функционирования пути поступления интактных белков посредством пиноцитоза даже с невысокой эффективностью в неонатальный период может быть необходимым для абсорбции достаточных количеств макромолекул с целью индукции антителообразования.
Ацидурия нейтральных аминокислот (болезнь Хартнупа)
Хорошо известно, что как ферментативные, так и транспортные функции клеток находятся под контролем соответствующих генов и, следовательно, зависят от опосредуемых мутационным процессом изменений. Примером повреждения процесса эпителиального транспорта аминокислот на генетическом уровне является болезнь Хартнупа, названная по имени исследователя впервые описавшего данное заболевание. Болезнь характеризуется неспособностью эпителиальных клеток кишечника всасывать и почек реабсорбировать нейтральные аминокислоты. В почках, где содержащиеся в плазме аминокислоты попадают в просвет проксимальных канальцев за счет ультрафильтрации, неспособность эпителиальных клеток реабсорбировать нейтральные аминокислоты проявляется в экскреции данных соединений с мочой (аминоацидурия). Дефект одной из транспортных систем эпителиальных клеток тонкого кишечника приводит к нарушению всасывания свободных аминокислот, происходящих из пищевых белков. Клинические признаки заболевания сходны с проявлениями, которые наблюдаются при существенном дефиците аминокислот (отрицательный азотистый баланс) и никотинамида (нарушения обмена аминокислот – реакции окислительного дезаминирования и нарушения обмена углеводов). В свою очередь, дефицит никотинамида определяется острой нехваткой триптофана, который является необходимым предшественником для биосинтеза этого важного компонента кофакторов NAD+ и NADP+.
Обследование пациентов с болезнью Хартнупа показало присутствие в плазматических мембранах энтероцитов функционирующих транспортных систем для ди- и трипептидов и наличие измененных транспортных систем аминокислот. Таким образом, вследствие того, что генетические дефекты компонентов систем транспорта нейтральных аминокислот ограничивают их поступление в организм – транспорт ди- и трипептидов остается практически единственным путем поступления продуктов расщепления пищевых белков в кровь (Silk D.B.A., Disorders of nitrogen absorption. Clinics in gastroenterology: Familial inherited abnormalities, Harries J.T. (ed). Vol 11: London: Saunders, 1982, 47-73).