

ПРИЛОЖЕНИЕ 7.1
Обмен 2,3-дифосфоглицерата
Одной из реакций гликолитического пути распада глюкозы является реакция превращения 3-фосфоглицерата в 2-фосфоглицерат. При этом фосфорильная группа 3-фосфоглицерата перемещается из третьего положения во второе с образованием 2-фосфоглицерата. Данную реакцию катализирует фосфоглицеромутаза. В целом мутазы представляют собой класс ферментов обеспечивающих внутримолекулярное перемещение химических групп как, например, в данном случае фосфорильной группы. Фосфоглицеромутазная реакция обладает замечательной особенностью: фосфорильная группа не просто перемещается от одного атома углерода к другому. Этот фермент зависит от присутствия в каталитических количествах 2,3-дифосфоглицерата, который поддерживает остаток His в активном центре фермента в фосфорилированном состоянии:
Е–His + 2,3-дифосфоглицерат ↔ Е–His–фосфат + 2-фосфоглицерат
Е–His–фосфат + 3-фосфоглицерат ↔ Е–His + 2,3-дифосфоглицерат
Сумма этих двух реакций дает нетто выражение мутазной реакции:
3-фосфоглицерат ↔ 2-фосфоглицерат
Анализ первой части реакции показывает, что мутаза действует как фосфатаза – фермент превращает 2,3-дифосфоглицерат в 2-фосфоглицерат. Однако при этом фосфорильная группа остается связанной с ферментом. Эта фосфорильная группа затем переносится на 3-фосфоглицерат, регенерируя 2,3-дифосфоглицерат. Поскольку фосфорилированная форма фермента относительно нестабильна и подвергается распаду до свободного фермента и неорганического фосфата очевидно должна существовать отдельная реакция, обеспечивающая генерирование 2,3-дифосфоглицерата, который будет рефосфорилировать любую молекулу свободного фермента.
Действительно, существует фермент – дифосфоглицератмутаза, который катализирует образование 2,3-дифосфоглицерата из 1,3- дифосфоглицерата и 3-фосфоглицерата. Особенно большое значение эта реакция приобретает в эритроцитах, поскольку 2,3-дифосфоглицерат служит аллостерическим регулятором транспортной функции гемоглобина человека. Весьма интересен механизм описываемой мутазной реакции. Обязательным ее участником является 3-фосфоглицерат, хотя в итоговой стехиометрии реакции он не фигурирует (рис. 7.1.1). Дифосфоглицератмутаза одновременно связывает 1,3-дифосфоглицерат и 3-фосфоглицерат. В составе
этого тройного комплекса происходит перенос фосфорильной группы от С-1 1,3-дифосфоглицерата на С-2 3-фосфоглицерата. В данной мутазной реакции 2,3-дифосфоглицерат выступает как мощный конкурентный ингибитор связывания 1,3-дифосфоглицерата. Таким образом, скорость синтеза 2,3- дифосфоглицерата отчасти зависит от его собственной концентрации.
Рис. 7.1.1 Схематическое изображение участия 3-фосфоглицерата в превращении 1,3-дифосфоглицерата в 2,3-дифосфоглицерат.
Другим регуляторным фактором реакции образования 2,3- дифосфоглицерата является концентрация 1,3-дифосфоглицерата, поскольку фермент не всегда насыщен этим соединением. В противоположность содержание 3-фосфоглицерата (в эритроцитах) почти всегда находится на уровне достаточном для насыщения мутазы. Отсюда следует, что скорость синтеза 2,3-дифосфоглицерата регулируется концентрациями несвязанных 1,3-дифосфоглицерата и 2,3-дифосфоглицерата. Избыточные количества 2,3- дифосфоглицерата гидролизуются под действием 2,3-дифосфоглицерат- фосфатазы с образованием 3-фосфоглицерата.
Итак, помимо того, что 2,3-дифосфоглицерат (2,3-DPG) является важным участником реакции превращения 3-фосфоглицерата в 2- фосфоглицерат в ходе гликолиза, этот органический фосфат служит принципиальным регулятором транспорта кислорода в эритроцитах. Он снижает сродство гемоглобина к кислороду, стабилизируя Т-состояние гемопротеида. Эритроциты характеризуются высокой концентрацией 2,3- дифосфоглицерата (обычно равной 4 мМ), что отличает их от других клеток, содержащих лишь следовые количества этого соединения (как, например, в гепатоцитах или миоцитах где 2,3-дифосфоглицерат используется лишь в качестве кофактора мутазной реакции).
Регуляторная функция 2,3-дифосфоглицерата в процессе транспорта кислорода реализуется благодаря способности данного соединения связываться с гемоглобином и, тем самым, влиять на сродство гемопротеида к кислороду. Действительно, в отсутствие 2,3-дифосфоглицерата Р50 для гемоглобина составляет 1 торр, тогда как в присутствии этого эффектора Р50 для гемоглобина возрастает до 26 торр. Это означает, что 2,3-DPG снижает сродство гемоглобина к кислороду в 26 раз. Физиологический смысл этого
явления сводится к следующему: в отсутствие 2,3-DPG гемоглобин в составе эритроцитов, проходя по капиллярам тканей, где рО2 равно ~26 торр, отдавал бы лишь небольшую часть связанного кислорода.
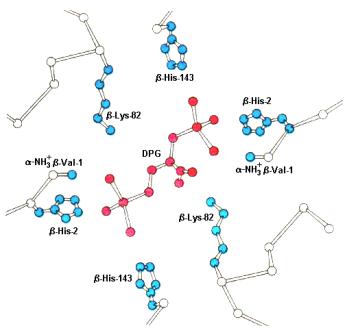
Методом ренгеноструктурного анализа было показано, что 2,3-DPG специфически связывается с дезоксигемоглобином в стехиометрии 1 молекула 2,3-DPG на 1 тетрамерную молекулу гемоглобина. (Arnone A., X-ray diffraction study of binding of 2,3-diphosphoglycerate to human deoxyhaemoglobin, Nature, 1972, 237, 5351, 146-149). Подобная стехиометрия взаимодействия эффектора с тетрамерами гемоглобина, структура которых описывается общей формулой α2β2, может указывать только на то, что 2,3- DPG связывается в центральной полости гемопротеида. Хорошо известно, что в гемоглобине человека HbA1 N-концевые последовательности β- субъединиц, обращенные в центральную полость тетрамерной молекулы белка, содержат общий кластер из восьми положительных зарядов,
представленных остатками β-Val-1, β-His-2, β-Lys-82, β-His-143 в каждой β-
цепи (рис. 7.1.2). Таким образом, связывание 2,3-DPG обусловлено образованием ион-парных взаимодействий между отрицательно заряженным
Рис. 7.1.2 Связывание 2,3-DPG с дезоксигемоглобином человека. 2,3-DPG электростатически взаимодействует с тремя положительно заряженными группами в каждой β-цепи белка. (Arnone A., X-ray diffraction study of binding of 2,3-diphosphoglycerate to human deoxyhaemoglobin, Nature, 1972, 237, 5351, 146-149).
эффектором, имеющим при физиологическом значении рН практически 4 отрицательных заряда и тремя положительно заряженными группами в каждой β-цепи белка.
В настоящее время стало понятным, почему 2,3-DPG снижает сродство гемоглобина к кислороду. Образуя перекрестные ион-парные взаимодействия с β-цепями, эффектор стабилизирует четвертичную структуру дезоксигемоглобина (так называемую Т-конформацию белка). Другими словами, 2,3-DPG сдвигает равновесие в сторону образования Т- конформации в ряду переходов Т↔R, где R-конформация соответствут полностью оксигенированному гемоглобину.
Клиническое значение 2,3-дифосфоглицерата
Выявление роли 2,3-DPG в транспорте кислорода позволило решить несколько проблем в разных областях клинической медицины. Так, например, на протяжении ряда лет оставалось непонятным, почему в крови, консервированной в среде цитрат – декстроза (общепринятая для консервирования крови среда), сродство гемоглобина к кислороду возрастает: вместо 26 торр значение Р50 становится равным 16 торр. В настоящее время установлено, что данное повышение сродства гемоглобина к кислороду обусловлено резким снижением содержания 2,3-DPG в эритроцитах с 4,5 до менее, чем 0,5 мМ за 10 дней хранения крови. Вместе с тем, сродство вливаемой консервированной крови к кислороду может иметь принципиальное значение в некоторых клинических случаях. Если больному переливают большое количество крови с высоким сродством к кислороду, то возникает опасность существенного снижения снабжения кислородом тканей. Попавшие в кровяное русло эритроциты, полностью лишенные 2,3- DPG, способны восстановить только половину его нормального содержания не менее чем за 24 часа. Такой длительный срок может оказаться критическим для тяжелых больных. Таким образом, в определенных условиях следует обращать внимание на то, чтобы при переливании крови больной получил эритроциты с нормальным сродством гемоглобина к кислороду. Простым добавлением 2,3-DPG нельзя повысить его содержание в эритроцитах, так как данный эффектор, обладая высоким зарядом, не способен проходить через клеточную мембрану. Можно, однако, предотвратить снижение концентрации 2,3-DPG в консервированных красных кровяных клетках, добавляя в среду консервирования инозин. Незаряженные молекулы инозина переносятся через мембрану эритроцита и внутри клетки в результате сложной серии реакций превращаются в 2,3-DPG. По этой причине в настоящее время инозин широко используется для сохранения функциональной полноценности консервированной крови.
Открытие роли 2,3-DPG позволило также понять и некоторые адаптивные механизмы, включающиеся при нарушении снабжения тканей кислородом. Возьмем для примера больных с тяжелой обструктивной эмфиземой легких. При этом заболевании затруднено поступление воздуха в бронхиолы; в результате артериальная кровь недостаточно насыщается