

случаях между альдегидом и ε-аминогруппами образуются простые Шиффовы основания или происходят только две из трех реакций, приведенных на рис. 5.1.1.
Поперечные связи, по-видимому, распределены не беспорядочно, а находятся в определенных положениях и часто располагаются в направлении концов мономеров коллагена. Остатки гистидина были обнаружены в полипептидных цепях только в положениях 89, 929 и 1034. Разнообразие и число поперечных связей варьируют при переходе от одного вида к другому.
Поперечные связи необычного характера были обнаружены в эластине. Три альдегидные группы взаимодействуют с одной аминогруппой лизина при помощи реакций альдольной конденсации, дегидратации и окисления, образуя десмозин и изодесмозин. В нативной молекуле эластина все четыре аминогруппы и карбоксильные группы участвуют, по-видимому, в образовании пептидной связи.
Генетические дефекты структуры коллагена
При изучении любого белка, играющего определенную роль в функционировании организма, исследователи постоянно сталкиваются с целым рядом генетических проблем. В случае коллагена возможность проявления вредных мутаций достаточно высока, поскольку этот белок кодируется больше чем одним набором генов. Действительно, было идентифицировано, по меньшей мере, четыре типа коллагена, характеризующихся вполне определенными различиями на молекулярном уровне, и было показано, что различные гены коллагена по-разному проявляют себя в разных тканях. Например, молекулы коллагена хрящей состоят преимущественно из трех идентичных α-цепей, отличающихся по аминокислотной последовательности от αl- и α2-цепей коллагена сухожилий и костей. Коллаген кожи маленьких детей и коллаген клапанов сердца и крупных артерий содержат полипептидные цепи двух других типов.
Четко выраженной наследственной аномалией коллагена является заболевание крупного рогатого скота дерматоспараксис, при котором кожа животного становится очень ломкой. Цепи коллагена при этом заболевании не обладают упорядоченной структурой и не способны, поэтому, образовывать фибриллы. Данная аномалия обусловлена дефектом проколлаген-пептидазы – фермента, отщепляющего пептиды с N-концов цепей коллагена.
Коллагеновым заболеванием человека является синдром Элерса– Данлоса, который в ряде случаев сопровождается деформацией суставов и искривлением позвоночника. Одна из форм этого заболевания обусловлена недостатком проколлаген-пептидазы, а другая – недостатком коллагена III. Содержание оксилизина в образующемся в данных условиях коллагене невелико, что препятствует образованию поперечных связей. Если в

организм животного с пищей попадают семена обычного сладкого горошка (Lathyris odoratus), то у них может наступить состояние латиризма. Поскольку люди также употребляют сладкий горошек в пищу, это состояние наблюдается и у людей; обычно оно приводит к деформации позвоночника и к разрыву аорты. Установлено, что биохимический эффект семян сладкого горошка обусловлен наличием в них β-цианаланина и продукта его декарбоксилирования β-амино-пропионитрила. Образование β-цианаланина у растений происходит в ходе пиридоксальфосфат-зависимой ферментативной реакции β-замещения из цистеина. Поэтому не удивительно, что в результате нарушений обмена серусодержащих аминокислот, описанных в лекции 5, наблюдаются также клинические проявления дефектов соединительной ткани.
Хотя точный механизм действия этих соединений неясен, есть основания полагать, что они являются ингибиторами лизил-оксидазы – фермента, необходимого для окисления аминогрупп боковых цепей лизина и оксилизина до альдегидных групп с последующим образованием поперечных связей в коллагене и в эластине. Аналогичного рода заболевание, передающееся по наследству, было обнаружено у мышей. Предполагается, что оно обусловлено или недостаточной активностью лизил-оксидазы, или дефектом в метаболизме меди. Кроме того, было высказано предположение, согласно которому одна из форм синдрома Элерса–Данлоса у людей вызвана недостаточной активностью лизил-оксидазы.
В настоящее время исследуется вопрос о влиянии аномалий в структуре коллагена на состояние суставов у человека. Было, например, показано, что при широко распространенном заболевании, остеоартрите, в хрящах вместо коллагена обычного типа [αl(II)]3 содержится коллаген, имеющий в своем составе α2-цепи, характеризующиеся пониженной степенью гликозилирования. С другой стороны, в хрупких костях людей, страдающих одним из видов наследственного заболевания, проявляемого в нарушениях процесса остеогенеза, наряду с содержащимся обычно в костях коллагеном типа I содержится также коллаген типа Ш.
Согласно имеющимся данным, содержание коллагена I в фиброзных атеросклеротических бляшках артерий человека повышено по сравнению с его содержанием в стенках нормальных артерий (содержащих в основном коллаген типа III).

ПРИЛОЖЕНИЕ 5.2
Полиамины и их физиологические функции
Роль орнитина в организме не ограничивается его участием в биосинтезе мочевины. Эта аминокислота вместе с метионином служит предшественником широко распространенных у млекопитающих (и бактерий) полиаминов – спермина и спермидина. В организме здорового человека синтезируется приблизительно 0,5 ммоль спермина в сутки. Фармакологические дозы полиаминов вызывают понижение температуры и снижение кровяного давления.
Спермидин и спермин (рис. 5.2.1) участвуют в различных физиологических процессах, общим признаком которых является связь с процессами пролиферации и роста клеток. Они являются факторами роста для культур клеток млекопитающих и бактерий и играют определенную роль в стабилизации интактных клеток, субклеточных органелл и мембран.
Рис. 5.2.1 Структура природных полиаминов. Обращает на себя внимание то, что спермидин и спермин являются полимерами диаминопропана и диаминобутана.
Благодаря тому, что молекулы полиаминов несут большое число положительных зарядов, они легко ассоциируют с полианионами, такими, как ДНК и РНК, и участвуют в таких фундаментальных процессах, как стимуляция биосинтеза ДНК и РНК, стабилизация ДНК и упаковка ДНК в бактериофагах. Полиамины оказывают влияние на синтез белка и являются ингибиторами целого ряда ферментов, включая такие важнейшие регуляторные белки как протеинкиназы.
Хотя в настоящее время не удается описать механизм действия полиаминов на специфические метаболические процессы, их важная роль в метаболизме млекопитающих не вызывает сомнения. Начальная реакция биосинтеза полиаминов катализируется орнитин-декарбоксилазой. В ходе этой реакции в результате декарбоксилирования орнитина (рис. 5.2.2) происходит образование путресцина (1,4-диаминобутана), являющегося основой для синтеза полиаминов.

Рис. 5.2.2 Начальная стадия биосинтеза полиаминов катализируется орнитиндекарбоксилазой. В результате декарбоксилирования орнитина происходит образование путресцина (1,4-диаминобутана), являющегося основой для синтеза полиаминов.
О важной физиологической роли данного фермента, единственной известной функцией которого является биосинтез полиаминов, свидетельствуют данные относительно того, что добавление в культуру клеток млекопитающих ингибиторов орнитин-декарбоксилазы (например, α- метилорнитина, или дифторметилорнитина) вызывает резкое усиление синтеза фермента.
Биосинтез полиаминов
На рис. 5.2.3 приведена схема метаболического пути биосинтеза полиаминов в тканях млекопитающих. Как следует из рисунка путресциновый фрагмент спермидина и спермина образуется из L-орнитина (интермедиата цикла мочевины), а диаминопропановый фрагмент – из L- метионина (с промежуточным образованием S-аденозилметионина). Орнитин-декарбоксилаза и S-аденозилметионин-декарбоксилаза являются индуцируемыми ферментами с коротким временем полужизни. Спермин- и спермидин-синтаза, напротив, не являются индуцируемыми и относительно «стабильны».
Из числа ферментов биосинтеза полиаминов у млекопитающих орнитин-декарбоксилаза и S-аденозилметионин-декарбоксилаза представляют интерес в плане их регуляции и возможности направленной химиотерапии. Время полужизни орнитин-декарбоксилазы (приблизительно 10 мин) меньше, чем у любого другого известного фермента млекопитающих; активность этого фермента очень быстро и повсеместно изменяется в ответ на различные стимулирующие воздействия. Например, добавление в культуру клеток млекопитающих гормона роста, кортикостероидов, тестостерона или фактора роста эпидермиса быстро повышает активность орнитин-декарбоксилазы в 10-200 раз. Было обнаружено, что при добавлении в культуру клеток полиаминов индуцируется синтез особого белкового антифермента (антизима), который, связываясь с орнитин-декарбоксилазой, ингибирует функцию фермента. Таким образом, активность орнитиндекарбоксилазы контролируется с помощью специфического белок-
белкового взаимодействия подобно регуляции активности трипсина белковым ингибитором трипсина. По отношению к орнитин-декарбоксилазе дифторметилорнитин можно рассматривать как «суицидальный ингибитор», т.е. соединение, превращающееся в ингибитор под действием самого фермента. Данное соединение используется для выделения штаммов мутантных клеток, характеризующихся гиперпродукцией орнитиндекарбоксилазы, а также в качестве ингибитора репликации клеток. Отсюда следует, что дифторметилорнитин действует как химиотерапевтический фактор, мишенью которого является орнитин-декарбоксилаза.
Рис. 5.2.3 Схема биосинтеза спермидина и спермина. Показаны исходные соединения, интермедиаты и ферменты, участвующие в синтезе полиаминов.
S-аденозилметионин-декарбоксилаза является единственной известной эукариотической декарбоксилазой, содержащей в качестве кофактора ковалентно связанный пируват (обычно декарбоксилазы содержат кофактор пиридоксальфосфат).
S-аденозилметионин-декарбоксилаза имеет сравнительно небольшое время полужизни (1-2 ч) и реагирует на действие стимуляторов клеточного роста подобно орнитин-декарбоксилазе, однако стимулирующий эффект
проявляется несколько медленнее. Активность S-аденозилметионин- декарбоксилазы ингибируется S-аденозилметилтиопропиламином (см. рис. 5.2.3) и стимулируется путресцином.
Катаболизм полиаминов
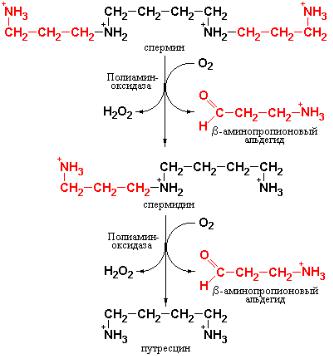
Катаболизм полиаминов в тканях млекопитающих осуществляется с участием фермента – полиамин-оксидазы, локализованного в пероксисомах гепатоцитов. Полиамин-оксидаза окисляет сначала спермин в спермидин и далее спермидин в путресцин (рис. 5.2.4). При этом диаминопропановые фрагменты полиаминов превращаются β-аминопропионовый альдегид.
Рис. 5.2.4 Схема катаболизма полиаминов. Пояснения в тексте.
Известно, что часть образующегося в ходе катаболизма полиаминов путресцина может окисляться до NH4+ и СО2, однако детали механизма этого процесса еще не выяснены. Основная же часть путресцина и спермидина выводится из организма в виде конъюгатов, главным образом, в форме ацетильных производных.