

Вегетативная, или автономная, нервная система
Адаптация, или приспособление организма к условиям существования, невозможна без соответствующих изменений в органах и системах, обеспечивающих обмен веществ и регуляцию всех процессов, происходящих в организме (рост.развитие, пищеварение, дыхание, выделение, кровообращение и т. д.). Взаимосвязь всех этих процессов и интенсивность их течения осуществляются вегетативным отделом нервной системы.
В зависимости от расположения вегетативных центров и структур, для которых предназначены 'нервные проводники, передающие импульсы от центров к исполнительным органам, вегетативный отдел нервной системы подразделяется на симпатическую й парасимпатическую части, из которых симпатическая часть — pars sympatica — предназначена для иннервации органов сердечно-сосудистой системы, а парасимпатическая — pars рага-sympatica — осуществляет иннервацию желез и всех внутренних органов. В связи с таким подразделением в отличие от цереброспинального отдела нервной системы вегетативный отдел имеет ряд существенных морфологи-ческих отличий.
Если эфферентные нервные волокна цереброспинальных нервов предназначены только для скелетных мышц, то в вегетативном отделе эфферентные волокна участвуют в иннервации не только мышц внутренних органов и желез внешней и внутренней ^секреции (парасимпатическая иннервация), но и всех отделов сердечно-сосудистой системы (симпатическая иннервация). Учитывая, что сосуды имеются во всех без исключения органах, то симпатические нервы и их волокна проходят как,к соматическим, так и к висцеральным органам. Поэтому 'каждый орган получает тройной, состав нервных волокон: чувствительных, двигательных (соматические иди парасимпатические) и симпатических.
Как и цереброспинальные, вегетативные нервы состоят из эфферентных и афферентных нервных волокон. Однако если афферентные нейроны цереброспинальных и симпатических нервов воспринимают раздражение как от соматических, так и от висцеральных органов, то в парасимпатической части афферентные нейроны имеют отношение лишь к висцеральным органам и располагаются в периферических ганглиях й интрамураль-ных сплетениях. Следовательно, парасимпатические центры получаюту информацию только со" стороны иннервируемых ими органов. Этим и объясняется относительная автономия вегетативного отдела нервной системы, имеющего характерные отличия в строении собственных рефлекторныхдуг. ' ■ ,
Все цереброспинальные нервные клетки лежат только в сером веществе спинного, или головного мозга и ганглиях их нервов, в то время как тела эфферентных клеток вегетативного отдела нервной системы находятся на периферии, а именно симпатические — в околопозвоночных или предпозвоночных ганглиях, а парасимпатические — в интрамуральных (внутристенных) или в экстрамуральных (органных) ганглиях органов го ловы, грудной,, брюшной и тазовой полостей. ,
Тела вставочных нейронов, передающих нервные импульсы с чувствительного звена на двигательное в вегетативных нервах, в противоположность спинномозговым образуют центры, расположенные или в боковых рогах спинного мозга (симпатическая часть), или в стволовой части головного и в крестцовом отделе спинного мозга (парасимпатическая часть), и своими отростками выходят за пределы мозга, заканчиваясь на телах эфферентных клеток, находящихся в периферических ганглиях. Поэтому в вегетативных нервах вставочные нейроны относятся к предузловым, или преганглионарным, нервным волокнам — neurofibra . preganglionica, а эфферентные — к постганглионарным — neurofibra postganglionica.
Наличие афферентной связи соматических и висцеральных органов через посредство преганглионарных (вставочных) нервных волокон с их эфферентными волокнами обусловливает их морфофункциональную зависимость как между собой, так и с высшими отделами головного мозга. Последнее не- только дает объяснение механизму выработки безусловных рефлексов со всех органов тела, действию ганглиолитиков и новокаиновых блокад, но и опровергает мнение об автономности вегетативного отдела нервной системы. Следовательно, вегетативный отдел нервной сиртемы представляет собой специализированную часть единой нервной системы, в которой все ее элементы подчинены высшим центрам, заложенным в коре головного мозга.
Развитие вегетативного отдела нервной системы. У беспозвоночных происходит выделение из соматического отдела нервной системы части нервных элементов, предназначенных для иннервации кишечной трубки. В частности, у аннелид есть нервы, которые отходят от глоточного ганглия и имеют в своем составе самостоятельные ганглии. У членистоногих эти нервы достигают значительного развития. Разделение вегетативного отдела на части проявляется-у высших насекомых, у которых краниальный й каудальные отделы соответствуют парасимпатической, а средний — симпатической .части.
У круглоротых самостоятельных вегетативных ганглиев нет, хотя блуждающий нерв и висцеральные ветви, отходящие от спинномозговых нервов, имеются. У высших хрящевых рыб происходит выделение сегментальных гцнглиев. Размещаются они метамерно вдоль аорты и соединяются ветвями со спинномозговыми нервами. От каждого ганглия отходят висцеральные ветви к внутренним органам, где они образуют сплетения с заложенными в них клеточными элементами. Начиная с костистых рыб, ганглии объединяются между собой межганглионарными связями, образуя правый и левый симпатические стволы: кранйально они заходят в область головы, а каудально — в область хвоста.
У земноводных за счет образования двойных связей между симпатическими ганглиями идёт образование двойного (поверхностного и глубокого) симпатического ствола. В связи с подразделением туловища на отделы у наземных позвоночных происходят, слияние отдельных. ганглиев между собой и соответствующие изменения во взаимоотношениях между поверхностным и глубоким симпатическими стволами, что особенно характерно для шейного отдела туловища.
В брюшном отделе у наземных позвоночных в'связи со значительным удлинением пищеварительной трубки, поворотами желудка и кишечника отмечают не-только сокращение числа сегментарных кровеносных сосудов, но и соответствующую концентрацию симпатических ганглиев.
В процессе эмбриогенеза у млекопитающих симпатическая часть вегетативного отдела нервной системы развивается из нейроэктодермы бокового отдела нервной трубки и выселяется за пределы спинного мозга по вентральным корешкам спинномозговых нервов. Образовавшийся парный симпатический-тяж затем фрагментируется на отдельные узлы симпатического ствола (паравертебральные узлы), имеющие соединения как между собой, так и с соответствующими спинномозговыми нервами и боковыми рогами сегментов спинного мозга. Часть нервных клеток смещается ближе к органам, образуя превертебральные ганглии или отдельные скопления нервных клеток по ходу висцеральных нервов.
Интрамуральные, парасимпатические ганглии образуются за счет вы-, селения клеточных элементов из стволовой части головного и крестцового отделов спинного мозга.
СОСТАВ ВЕГЕТАТИВНОГО ОТДЕЛА НЕРВНОЙ СИСТЕМЫ
Вегетативный отдел нервной системы включает вегетативные центры, вегетативные ганглии, вегетативные нервы и вегетативные сплетения.
Вегетативные центры заложены вокруг мозговых желудочков, моз-гового водопровода и центрального спинномозгового канала в виде скопле-ний серого вещества (одиночные или конгломераты ядер). Их можно под-разделить: а) на центры ^гипоталамуса (сосудосуживающие — симпатиче-ские, сосудорасширяющие и нейросекреторные — парасимпатические цент-ры); б) мозгового ствола, в котором выделяются центры среднего мозга(зрачковое, или ядро Эдингер — Вестфаля, — парасимпатические центры,ведающие функцией цилеарных и зрачковых мышц), центры мозгового мос-та (ростральное слюноотделительное и слизисто-слезноносовое парасимпа-тическое ядра), центры продолговатого мозга (каудальное слюноотделитель-ное ядро и ядра блуждающего нерва); в) спинного мозга (симпатические ядрапромежуточнолатерального тракта или спинномозговой центр Якобсона,располагающийся в 'пределах от С VIII до L II — L IV, и парасимпати-ческие центры крестцового отдела, находящегося на уровне L IV — L Vи даже Co II). , ' . _ - ■
Вегетативные ганглии подразделяются на пара- и превертебральные, органные и внутристенные.
а) паравертебральные располагаются метамерно с правой и левой"сторон позвоночного столба и составляют вместе с межганглионарными вет-вями основу симпатического ствола. Паравертебральные ганглии соединеныс соответствующими спинномозговыми нервами преганглионарными, белы-ми (rr. communicans alba) и серыми (гг. communicans grisea) соединитель-ными ветвями. Первые выходят из промежуточнолатерального тракта всоставе двигательного корешка и соединяют центр с соответствующим ган-глием; от последнего берут начало постганглионарные волокна, направляю-щиеся к внутренним органам или возвращающиеся (rr. communicans grisea)в состав спинномозгового нерва, с ветвями которого принимают участие виннервации соматических органов;
б) превертебральные ганглии находятся в отдалении от позвоночногостолба и располагаются в местах отхождения крупных артериальных со-судов (наружная и внутренняя сонные, плечеголовная, чревная, краниаль-ная и каудальная брыжеечные), участвуют в образовании соответствующих'нервных сплетений;в) органные и внутристенные (интрамуральные) ганглии находятсявблизи органа или внутри его стенки, Они небольших размеров, распола-гаются по ходу отдельных нервов, их ветвей (ресничный, крылонёбный,нижнечелюстной, ушной, подъязычный) или входят, в состав органныхсплетений (pi. myentericus, pi. subserosus, pi. pelvinus). '
Вегетативные эфферентные нервные волокна, отходящие от центров,и афферентные нервные волокна, идущие от органов в центры или соответ-ствующие вегетативные ганглии, могут образовывать вегетативные нервы(внутренностные нервы симпатического ствола) или входить в состав че-репных и спинномозговых нервов. *
Вегетативные сплетения образуются нли по ходу кровеносных сосудов, которые они сопровождают, или в воротах органа, или непосредственно внутри его стенки.
Отличия симпатической и парасимпатической частей вегетативного отдела нервной системы заключаются в следующем:
центры симпатической части в своем расположении имеют зональный характер (промежуточнолатеральный тракт спинного мозга от С VIII до L IV),. парасимпатические — очаговый (средний, продолговатый мозг и крестцовый отдел спинного мозга); >
афферентные нейроны в симпатической части могут заканчиваться как в ганглиях, так и в центрах, в парасимпатической части — только в центрах; , .
вставочные нейроны в симпатической части имеют сравнительно короткие аксоны, которые заканчиваются в пара- или в превертеОральных ганглиях, тогда как в парасимпатических они длинные и заканчиваются или в органных (ресничный, крылонёбный, подъязычный), или в интра-ррганных узлах; 4
v
1
1
эфферентные волокна в симпатической-части длинные, а в парасимпатической — короткие.
СИМПАТИЧЕСКАЯ ЧАСТЬ ВЕГЕТАТИВНОГО ОТДЕЛА НЕРВНОЙ СИСТЕМЫ
. В состав симпатической части — pars sympathicus —
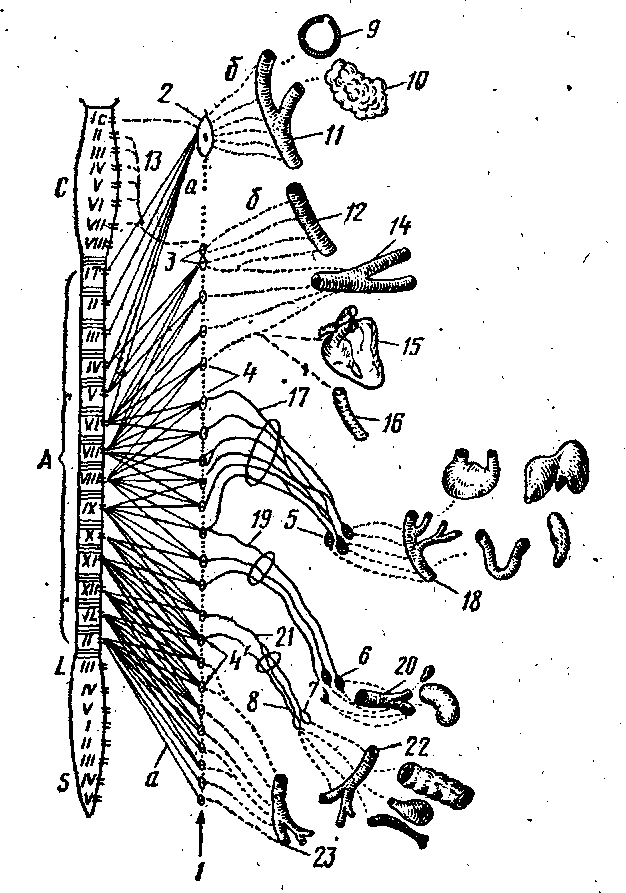
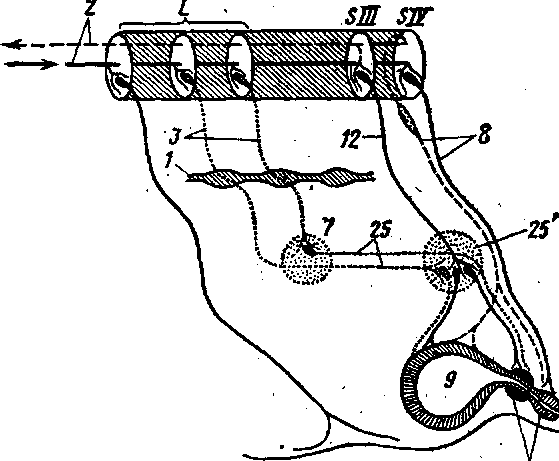
Рис. 274. Схема строения симпатической части вегетативного отдела нервной системы: А — грудопоясничиый, отдел спинного мозга; С — шейные; L — поясничные и S— крестцовые спинномозговые сегменты; 1 — симпатический ствол; 2 — краниальный шейный ганглий; 3 — звездчатый ганглий; 4 — грудные ганглии; 5 — чревное сплетение; 6 — почечное и надпочечное . сплетения; 7 — каудаль-ное брыжеечное сплетение; 8 — подчревное сплетение; 9 — в сосуды глаза; 10 — в сосуды желез; 11 — в сосуды головы; 12 — в сосуды шеи; 13 — позвоночный нерв; 14 — в сосуды грудной конечности; 15 — в/ сердце; 16 — в сосуды легких; 17 — большой внутренностный нерв; 18 — в сосуды желудка, печени, селезенки и кишечника; 19 — малый- внутренностный нерв; 20 — в сосуды надпочечников и почек; 21 — под-чревный нерв; 22 — в сосуды толстого отдела кишечника, мочеполовых органов; 23 — в сосуды тазовой конечности; а — пре-ганглионарные и б — постганглионарные нервные волокна.
входят центры, заложенные в боковых рогах спинного мозга (промежу-точнолатеральный тракт) в пределах от С VIII до LII—IV(SI), симпатический ствол с паравертебральными ганглиями и внутренностными нервами, участвующими вместе с ветвями блуждающего и тазовых нервов в образовании сплетений (рис. 274, цв^ табл. XVI).
Симпатический ствол — truncus sympathicus
(рис.
274—/) — парный (dexter
et sinister), проходит
вдоль позвоночного столба и подразделяется
на шейный, грудной, поясничный, крестцовый
и хвостовой отделы. Правый и левый
симпатические стволы имеют в своем
составе симпатические ганглии — gn.
trunci sympathici, соединенные
между собой межганглионарными ветвями
— rr.
iriterganglionares —
и с соответствующим спинномозговым
нервом соединительными ветвями — гг.
сопл-municantes
(а).
Число
симпатических ганглиев не во всех
участках тела соответствует числу
нейросегментов, так как они могут*
сливаться между


собой;
например, в шейном Отделе их три, в
грудном отделе первые три, сливаясь
между собой и с последним шейным,
образуют шейногруд-ной (звездчатый)
узел — gn.
cervicothoracicum (stellatum); в
крестцовом отделе они также нередко,
особенно у4
плотоядных и птиц, сливаются в один
общий узел. Могут быть и Дополнительные
симпатические ганглии, которые чаще
располагаются в межганглио-нарных
ветвях, — промежуточные ганглии —
gn.
intermedia. По
ходу симпатические стволы правой и
левой сторон иногда в грудном и
поясничном отделе имеют поперечные
связи, а в области последнего крестцового
сегмента соединяются в общий ствол;
на месте соединения отмечают непарный
симпатический ганглий — gn.
impar. От
последнего на хвостовой отдел
продолжается непарный симпатический
ствол, насчитывающий в своем составе
2—4 хвостовых ганглия — gn.
caudalia.
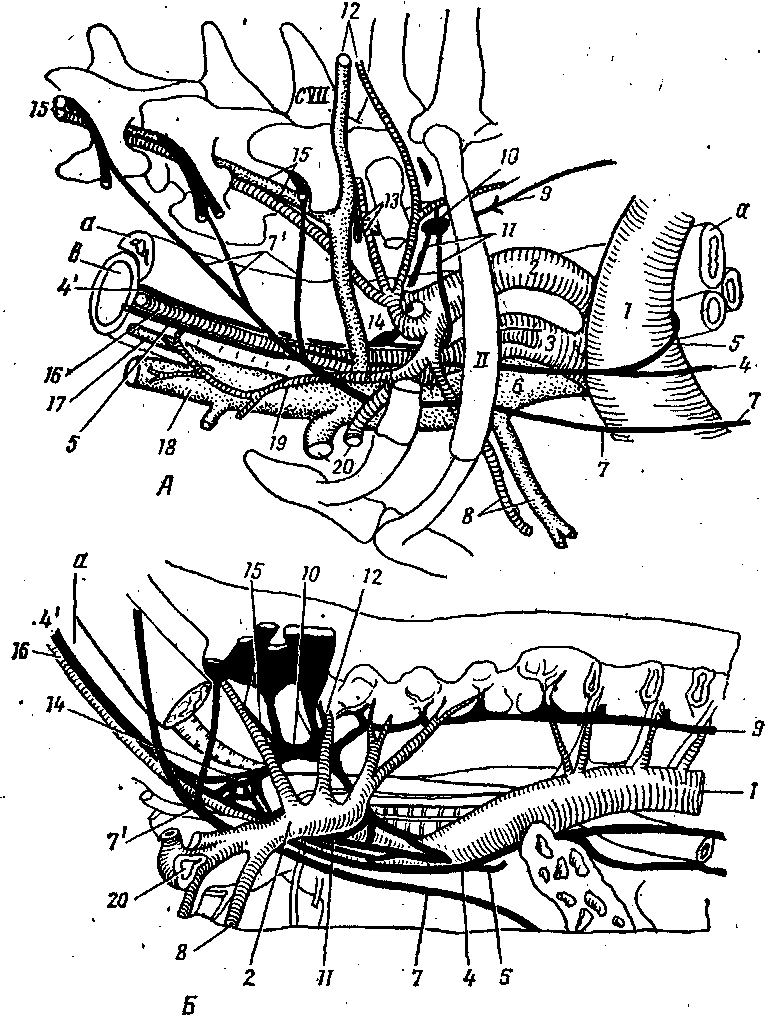
Шейный отдел симпатического ствола .берет начало от звездчатого узла двумя ветвями (рис. 275—Л), из которых дистальная ветвь снизу охватывает подключичную артерию, образуя подключичную петлю, а затем, соединившись с проксимальной ветвью, продолжается как truncus sympathicus. На месте соединения проксимальной и дистальной ветвей в общий симпатический ствол находится средний шейный ганглий — gn. cervicale media (14). Шейный симпатический ствол, направляясь к краниальному шейному узлу, тесно прилежит к блуждающему нерву, образуя с ним общий вагосимпатиче-ский ствол — truncus vagosympathicus (4'). Последний проходит в желобе, образованном пищеводом и трахеей вместе с общей сонной .артерией, находясь с ней в одном
общем соединительнотканном футляре. По ходу от симпатического ствола отходят ветви к блуждающему нерву и к общей сонной артерии, вокруг которой они образуют общее сонное сплетение — pi. caroticus communis. От него отделяется ряд веточек к гортани и глотке (rr. laryngopharyngei). Кранйально шейный симпатический ствол, обособившись от блуждающего нерЕа, вступает в краниальный шейный ганглий — gn. cervicale craniele (рис. 274—2).
От среднего шейного ганглия отходят постганглионарные волокна ь составе шейного сердечного нерва — п. cardiacus cervicalis, участвующего вместе с сердечными ветвями от звездчатого узла и блуждающего нерва в образовании сердечного сплетения — pi. cardiacus.
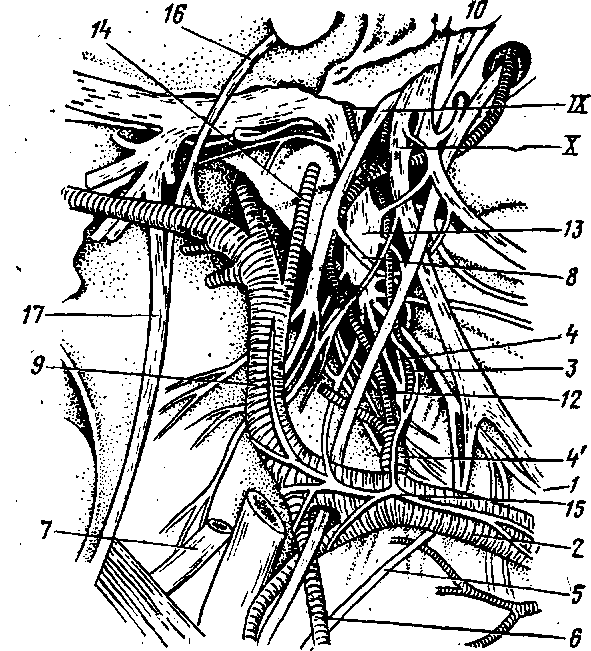
Краниальный шейный ганглий — gn. cervicale craniale (рис. 274 , 276, 277—13) — крупных размеров, располагается на дорсомедиальной поверхности внутренней сонной артерии и отдает от себя постганглионарные ветви, проходящие в составе яремного, внутреннего и наружных сонных нервов.
Внутренний сонный нерв — п. caroticus internus — образует внутреннее сонное сплетение — pi. caroticus internus, которое сопровождает все разветвления внутренней сонной артерии. На пещеристом синусе мозговых оболочек он формирует pi. cavernosus. Далее направляется в гипофиз и на черепномозговые нервы, а через них уже достигает сосудов радужной оболочки и слезной железы. Из внутреннего сонного сплетения выходит большой глубокий каменистый нерв — п. petrosus profundus major. Он проникает через видиев канал.в крылонёбный.узел на одноименном керве и в составе последнего идет в сосуды органов глазницы и слизистой оболочки носа и ротовой нолости.
Яремный нерв — п. jugularis — одной ветвью идет в gn. proximale (jugularis) вагуса и в ствол вагуса, а другой' ветвью — в gn. distale (petro-sum) язычногдоточного нерва и в ствол язычноглоточного нерва. Волокна яремного нерва вступают в, сосуды глотки, пищевода, гортани и трахеи.
Наружные сонные нервы — nn. carotici externi — проходят по ветвям одноименной артерии, образуя наружное сонное сплетение — pi. caroticus externus.
Соединительные ветви — nn. communicantes к IX, X и XII черепно-мозговым нервам, к 1-му шейному нерву и одну веточку в glomus caroticum,-располагающегося на внутренней сонной артерии.
. Грудной отдел симпатического ствола— pars thoracica (цв. табл. XVI— 12) — берет начало от шейногрудного, или. звездчатого ганглия (13) и, направляясь каудально, проходит через ножки диафрагмы в брюшную полость. От звездчатого узла отходит несколько нервов. '
а) Позвоночный нерв — п. vertebralis (15) — вступает в поперечноеотверстие 6-го шейного позвонка и проходит в поперечном канале до 2-гашейного позвонка, отдавая по своему ходу ветви к шейным спинномозго-вым нервам и образуя вокруг позвоночной артерии позвоночное сплетение —pi. vertebralis. Постганглионарные волокна в, составе ветвей спинномозго-вых нервов иннервируют сосуды шеи и спинного мозга.
б) Rami communicantes к С VIII, Th I—Th II участвуют в образовании,плечевого сплетения.
в) Шейные сердечные нервы — nn. cardiaci cervicales — в количестветрех направляются в сердечное сплетение. От грудных ганглиев — gn.thoracica — отходят: грудные сердечные нервы, участвующие в образо-4вании сердечного ф1. cardiacus) и аортального сплетений (pi. aorticus thora-cicus), легочные ветви — гг. pulmonales, образующие легочное сплетение(pi. pulmonalis). Кроме того, начиная с 6-го грудного сегмента, часть пре-ганглионарных волокон образует ветви, бтходящие от симпатического ство-ла и формирующие большой внутренностный нерв — п. splanchnicos major(рис. 274-/7). На месте соединения ветвей, образующих внутренностныйнерв, располагается внутренностный ганглий -г- gn. splanehnicum, от ко-торого нерв направляется в чревные ганглии — gn. celiaca (рис. 278—5).

.
Рис. 278. Вегетативные сплетения брюшной
полости: 1—
truncus
sympathicus; 4
—
rr.
communicantes; 5
—
plexus
ce-liacus; €
— plexus
renalis et plexus intermesentericus; 7
— plexus
mesentericus caudalis; 9
—
truncus
vagalis dorsalis et rr. celiaci; 17
—
n.
splanchnic.us major; 25
—
n.
hypogastrics; a
—
аорта; в
—
чревная
артерия.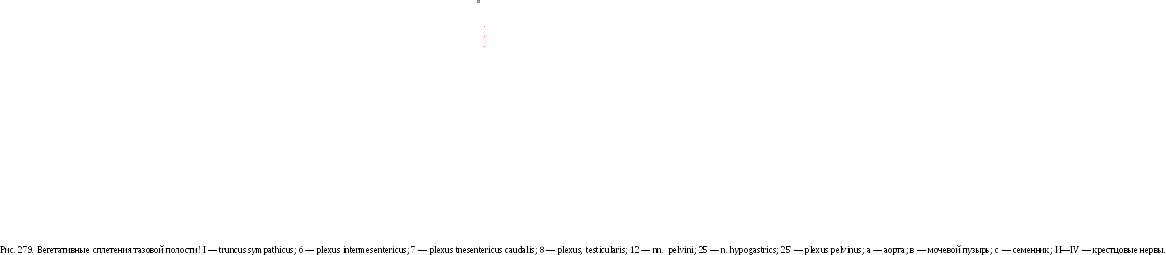
На уровне последних двух грудных-ганглиев от симпатического ствола»отходят малый внутренностный нерв — п. splanchnicus minor, вступаю-щий в краниальный брыжеечный узел — gn. mesentericus cranialis, и самыйзадний внутренностный нерв, или почечная ветвь — п. splanchnicus imus,s.r. renalis, направляющаяся к почке, где вступают в почечные ганглии —gn. renalia. "
Поясничный отдел симпатического ствола имеет поясничные ганглии — gn. lumbalia, от которых' отходят поясничные внутренностные нер: вы — nn.' splanchnici lumbales, вступающие в каудальный брыжеечный ганглий'— gn. mesentericus caudalis (рис. 278). '
Крестцовый отдел симпатического ствола служит продолжением поясничного и имеет в своем составе симпатические ганглии, которые могут объединяться между собой. От них отходят ветви, образующие крестцовые внутренностные нервы — пн. splanchnici sacrales, вступающие в тазовые ганглии—gn.pelvina. Каудально правый и левый симпатические стволы объединяются, образуя непарный симпатический ствол. На месте соединения симпатических стволов располагается непарный ганглий — gn. impar. Продолжающаяся хвостовая часть симпатического ствола имеет 2 —4 хвостовых ганглия — gn. caudalia, соединяющихся с Хвостовыми спинномозговыми нервами (рис. 279).
ПАРАСИМПАТИЧЕСКАЯ ЧАСТЬ ВЕГЕТАТИВНОГО ОТДЕЛА НЕРВНОЙ СИСТЕМЫ
Парасимпатическая часть — pars parasympathica, как и симпатическая, имеет: а) центры, расположенные в стволовом отделе головного мозга (сред-немозговая и продолговатомозговая Части) и крестцовом отделе спинного мозга; б) ганглии, находящиеся или вблизи органа, или непосредственно в его стенке, или по ходу нерва, и в) нервные волокна (преганглионарные ■ и постганглионарные), проходящие в составе как черепных, так и спинномозговых нервов. Последние вместе с ганглиями и симпатическими нервами участвуют в образовании вегетативных (автономных) сплетений.
ЦЕНТРЫ ПАРАСИМПАТИЧЕСКОЙ ЧАСТИ ВЕГЕТАТИВНОГО ОТДЕЛА'НЕРВНОЙ СИСТЕМЫ, ИХ НЕРВНЫЕ ПРОВОДНИКИ И ГАНГЛИИ
Среднемозговая часть представлена парасимпатическим , ядром глазбдвигательного нерва — nuclei parasympathici п. oculomotorii (цв. табл. XV—/), расположенного на дне мозгового водопровода вблизи передних бугров четверохолмия, медиально от двигательного ядра глазодвигательного нерва. Выйдя в составе'глазодвигательного нерва, преганглионарные волокна заканчиваются на эфферентных клетках ресничного узла— gn. ciliare (2), отростки которых образуют короткие цилиарные нервы — nn. ciliares. Последние, соединяясь с ветвями глазничного и тройничного нервов, вступают в глазное яблоко и разветвляются в ресничной мышце и сфинктере зрачка (<?).
Продолговатомозговая часть включает три пары ядер; от них берут начало преганглионарные волокна, входящие в состав лицевого, языко-глоточного и блуждающего нервов.
а) В составе лицевого нерва проходят чувствительные, двигательные и парасимпатические (слюноотделительны^ путь) нервные волокна. Последние берут начало от рострального слюноотделительного ядра —»- nucleus salvatorius rostralis (VII), расположенного в мозговом мосту на дне переднего отдела ромбовидной ямки. По выходе из мозга преганглионарные парасимпатические волокна отделяются от лицевого Нерва и образуют chorda tympani (8), которая присоединяется к язычному нерву и через, соединительную ветвь достигает нижнечелюстного ганглия — gn. mahdibulare (9),
Рис. 280. Иннервация околоушной железы:
А — у собаки; Б — у свиньи (по Г. П. Ме-лехину); 1 — околоушная железа; 2 —. нижнечелюстная железа; 3 — яремная веяв; 3' — верхнечелюстная вена; 4, 4' — plexus parotides; 5 — ductus parotideus; 6 — ganglion oticum.
а у плотоядных и подъязычного — gn. sublinguale. От последних часть постганглионарных волокон в составе железистых ветвей разветвляется в нижнечелюстной и подъязычной железах.
Другая часть постганглионарных волокон, проходящих в составе лицевого нерва, берет начало от слезного ядра— nucleus lacrimalis," расположенного рядом с предыдущим. Отходящие от него волокна относятся к слезоотделительному пути и, отделившись от лицевого нерва, входят в состав большого каменистого нерва — п. petrosus major (4), который входит в крылонёбный ганглий-~-gn. pterygopalatinum (5). От последнего через соединительные ветви сначала к верхнечелюстному, а через него — к слезному нерву (ветвь глазничного нерва) постганглионарные волокна проходят в слезную железу (6). Кроме того, часть волокон Через соединительные ветви к каудальному носовому, большому и малому нёбным нервам (ветви крылонёбнбго нерва) иннервирует слизистую оболочку носовой полости (7), твердбго и мягкого нёба.
б) В составе языкоглоточного нерва проходят чувствительные, дви-гательные и парасимпатические (преганглионарные) Волокна. Последниеберут начало от каудальндго слюноотделительного ядра — nucleus salvato-rius caudalis (IX). Эти волокна от языкоглотрчного нерва вступают в бара-банный нерв — п. tympanicus (//), а через него — в ушной ганглий — gn.oticum (12), расположенный на нижнечелюстном или крыловом нерве иимеющий соединительные веточки с барабанной струной и симпатическимсплетением среднемозговой артерии. От ушного узла отходят постганглио-нарные волокна в составе железистых ветвей к околоушной (13') и щечным(13) железам (рис. 280).
в) В составе блуждающего нерва проходят разнообразные нервныепроводники, представляющие афсрерентные и эфсрерентные парасимпатиче-ские, чувствительные и двигательные соматические и, наконец, эфгрерент-ные симпатические волокна. Такое разнообразие состава нервных волоконобусловило то, что блуждающий Нерв берет начало от трех ядер: от двой-ного — nucleus ambiguus (nucleus motorius n. vagi), парасимпатического —
nucleus parasympathicus n. vagi, расположенного поверхностно в каудальной части ромбовидной ямки, и от ядра одиночного тракта — nucleus tractus solitarii. По выходе" из продолговатого мозга блуждающий нерв имеет два ганглия: проксимальный (яремный) — gn. proximale — содержит в основном чувствительные клетки (соматические и парасимпатические), обеспечивающие связь периферических органов с центрами, заложенными в цродолго-ватом мозге; дистальный (узловатый) — gn. distale — прилежит к краниальному шейному ганглию, соединяется с ним мелкими веточками, содержит эфферентные соматические и вегетативные нервные волокна, направляющиеся в составе соединительных ветвей к подъязычному и добавочному нервам, а также глоточную ветвь в мышцы глотки и начальную часть пищевода и краниальный гортанный нерв для гортани, щитовидной железы и в глоточное сплетение. Продолжающийся блуждающий нерв смешанный, содержит в своем составе пучки волокон, составляющих основу возвратного и дёпрессорного нерва (см. Черепномозговые нервы — X пара). Вступив в грудную полость и отделившись'от симпатического ствола, блуждающий нерв продолжается в каудальном направлении, принимая участие в образовании сплетений органов грудной и брюшной полостей.
Крестцовая часть представлена парасимпатическими крестцовыми ядрами — nuclei parasympatici sacrales, располагающимися в боковых рогах серогб вещества последних крестцовых нейросегментов. Преганглионарные парасимпатические волокна, выйдя в составе вентральных корешков из спинного мозга, объединяются в 1—3 тазовые нервы — nn. pelvini, которые проходят по латеральной поверхности прямой кишки. Разделившись на множество веточек, тазовые нервы вместе с ветвями подчревного нерва участвуют в образовании тазового- сплетения.
ВЕГЕТАТИВНЫЕ (АВТОНОМНЫЕ, ИЛИ ВИСЦЕРАЛЬНЫЕ) СПЛЕТЕНИЯ
Грудное аортальное сплетение — pi. aorticus thoracicus — включает как непосредственно сплетение, располагающееся по ходу грудной аорты, так и отдельные органные сплетения, которые" образуются постганглио-нарными ветвями, отходящими как от узлов грудного симпатического ствола, так и от блуждающих нервов.
а) Сердечное сплетение — pi. cardiacus — образуется сердечными нер-вами, отходящими от среднего шейного, звездчатого и грудных ганглиев,а также сердечными ветвями блуждающего, депрессорным нервом, ветвямивозвратного, каудального гортанного нервов. В своем составе сплетение име-ет различной величины сердечные ганглии — gn. cardiaca. Сплетение под-разделяется на поверхностное, расположенное между дугой аорты и легоч-ной артерией, и глубокое, находящееся позади дуги аорты.
б) Легочное сплетение — pi. pulmonalis — образовано легочными вет-вями грудного симпатического ствола и бронхиальными ветвями блуждаю-щих нервов. в) Пищеводное сплетение — plexus esophageus — образуется дор-сальными и вентральными стволами блуждающего нерва. а
Брюшное аортальное сплетение—plexus aorticus abdominalis — окружает брюшную аорту и ее ветви. Оно подразделяется на чревное, краниальное и каудальное брыжеечное и тазовое сплетения.
а) Чревное сплетение — pi. celiacus (7) — образовано большим внутренностным нервом и чревными ветвями дорсального ствола вагуса. Центром чревного сплетения служит парный чревный ганглий — gn. celiacum (рис. 278—5), имеющий полулунную форму и располагающийся вокруг чревной артерии. От него отходят ветви, сопровождающие сосуды чревной артерии и участвующие в образовании органных сплетений: желудочного — pi. gastrici (у жвачных — pi. ruminalis dexter et sinister, pi. reticularis), селезеночного — pi. lienalis, печеночного — pi. hepaticus. От же
лудочного
сплетения отходят ветви, участвующие
также в образовании сплетений
двенадцатиперстной кишки, поджелудочной
железы, печени.
б) Краниальное брыжеечное сплетение — plexus mesentericum craniale — образовано ветвями малого внутренностного нерва при участии большого внутренностного и ветвей блуждающего нерва. Его центром служит краниальный брыжеечный ганглий, который располагается с каудальной стороны краниальной брыжеечной артерии и имеет комиссуральные связи с чревными узлами. Отходящие от него ветви частично направляются по краниальной брыжеечной артерии, образуя на ней одноименное сплетение, а частью волокон направляются к каудальному брыжеечному узлу, образуя межбрыжеечное сплетение — plexus intermesentericus. Вместе с ветвями чревного сплетения краниальное брыжеечное сплетение участвует в образовании сплетения поджелудочной железы, надпочечников, а с ветвями от поясничного отдела симпатического ствола — почечного, мочеточ-никового, яичникового (у самцов — семенникового). К. последним могут подходить и ветви от каудального брыжеечного сплетения (С- И. Шведов, 1977). В этих сплетениях могут быть более мелкие ганглии — gn. aortico-renalis, gn. renalia.
На кишечнике ветви краниального брыжеечного вместе с ветвями каудального брыжеечного сплетения образуют кишечное сплетение — pi. entericus, которое подразделяется на подсерозное, мышечно-кишечное и подслизистое — pi. subserosa, myentericus et submucosus.
в) Каудальное брыжеечное сплетение — plexus mesentericus- caudalis (рис. 279—7) — образуется внутренностными нервами поясничного отдела. От каудального брыжеечного узла отходят ветви, образующие ободочное сплетение — pi. colicus, краниальное ректальное сплетение — pi. rectalis cranialis —г и подчревньщ нерв — п. hypogastrics (25), направляющийся в-тазовую полость, где вступает в тазовое сплетение. Брюшноаортальное сплетение в каудальном направлении продолжается в подвздошное сплетение — pi. iliaci, от которого часть ветвей проходит по наружной подвздошной артерии и образует бедренное сплетение — pi. femoralis.
Тазовое сплетение — plexus pelvinus (25') — образуется тазовыми нерваМи — nn. pelvini (12), отходящими от крестцового отдела спинного мозга, крестцовой части симпатического ствола к подчревным нервам. В центре сплетения находятся тазовые ганглии — gn. pelvini.
Ветви, отходящие от тазового сплетения, образуют органные сплетения: среднее и каудальное ректальное—pi. rectalis medii et caudalis, предстательное — pi. prostaticus, семявыносЯщего протока — pi. deferentalis (у самок — маточновлагалищное — pi. uterovaginalis), пузырное — pi. vesicalis (рис. 281). От тазрвого сплетения отходят нервы в кавернозное тело
пениса — nn. corporis cavernosi penis (у самок — nn. corporis cavernosi clitoridis).
Таким образом, тазовое сплетение представляет собой сложное образование, включающее в свой состав как парасимпатические, так и симпатические нервные волокна. Отходящие от него ветви к внутренним органам тазовой полости образуют внутристенные органные сплетения, где наряду с нервными волокнами имеется значительное число нервных клеток и их скоплений в виде узелков различной величины. Через подчревный нерв тазовое сплетение морфологически и функционально объединяется с каудальный брыжеечным^ с плетением, а через соединительные ветви — й со спинальны-ми нервами, предназначенными для иннервации области промежности, анального отверстия и наружных половых органов.
Двойная (симпатическая и парасимпатическая) иннервация органов тазовой полости, включая наружные половые органы, с одной стороны, обусловливает их тесную морфофункциональную зависимость. За счет этих связей происходит координация функций спинально-анальных и генитоспиналь-ных центров с сосудорасширяющими центрами, заложенными вдоль центрального спинномозгового канала. Это ставит в зависимость .осуществление таких рефлексов, как мочеиспускание, дефекация, эрекция и эякуляция. С другой стороны, морфологические взаимосвязи тазового сплетения с каудальный брыжеечйым (через подчревный нерв), а последнего с краниальным брыжеечным (через брюшноаортальное сплетение и межбрыжеечный тракт) морфофункционально объединяют все отделы желудочно-кишечного тракта как между собой (Г. И. Яшина, 1983), так и с органами мочеполо-вого аппарата.
ОРГАНЫ ЧУВСТВ-ORGANA SENSUUM
Анализатор — analisator, по И. П. Павлову, — это «сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозге». Анализаторы, являющиеся обязательной частью нервной системы, состоят из: а) рецепторов; б) периферических проводящих путей и в) мозгового отдела с подкорковыми, и корковыми центрами, соединенными центральными проводящими путями.
Рецепторы — receptores — относятся к периферическому аппарату анализаторов, которые воспринимают и трансформируют, физико-химическую энергию внешней и внутренней среды в нервное возбуждение. По периферическим проводящим путям анализаторов, входящих в состав спинномозговых и головномозговых нервов, нервное возбуждение передается от рецепторов в подкорковые и корковые чувствительные центры. В коре большого мозга, которая представляет «совокупность корковых концов анализаторов», происходят анализ и синтез воспринятых раздражений, на основании чего и возникает ответная реакция. Различают экстеро- и интерорецептбры.
Экстерорецепторы воспринимают различные раздражения, исходящие непосредственно из внешней среды: химические раздражения воспринимаются вкусовыми и обонятельными рецепторами; физические раздражения — зрительными, слуховыми и равновесными рецепторами, терморецепторами и др.;~ механические раздражения — осязательными (тактильными) рецепторами. Отличительная черта экстербрецепторов заключается в том, что все возникающие в них возбуждения, достигнув коры мозга, становятся уже ощущениями, т. е. осознаются. В силу этого экстерорецепторы издавна получили ошибочное название «органы чувств» — organa sen-suum, тогда как «чувства» возникают в коре большого мозга, а не в самих рецепторах. Ощущение есть превращение энергии внешнего раздражения в факт сознания. Сознание — это функция коры большого мозга.
Интерорецепторы воспринимают раздражения, возникающие в различных органах и тканях самого организма вследствие изменений, происходящих в их деятельности и состоянии как в норме, так и в патологии. Сигнализация в ЦНС (центральную нервную систему) с интерорецеп-торов обеспечивает нормальный обмен веществ, местную регуляцию кровоснабжения тканей и органов, координацию функций как различных систем органов, так и внутри каждой системы. Интерорецепторы находятся даже в ганглиях симпатической нервной системы. Например, усиление мышечной работы при движении вызывает необходимость в повышенной доставке питательных веществ и кислорода в работающие мышцы. Это достигается более интенсивным функционированием органов дыхания и дыхательных мышц, сердечно-сосудистой системы, органов пищеварения, терморегуляции и выделения.
Хотя в норме все импульсы с внутренних органов не доходят до сознания, тем не менее они создают общий фон для нервной деятельности в це-
лом,
что вызывает у человека чувство или
удовлетворения, или неудовлетворения,
или недомогания наряду с такими общими
чувствами, как голод, жажда, половое
влечение, усталость или, напротив, позыв
к деятельности.
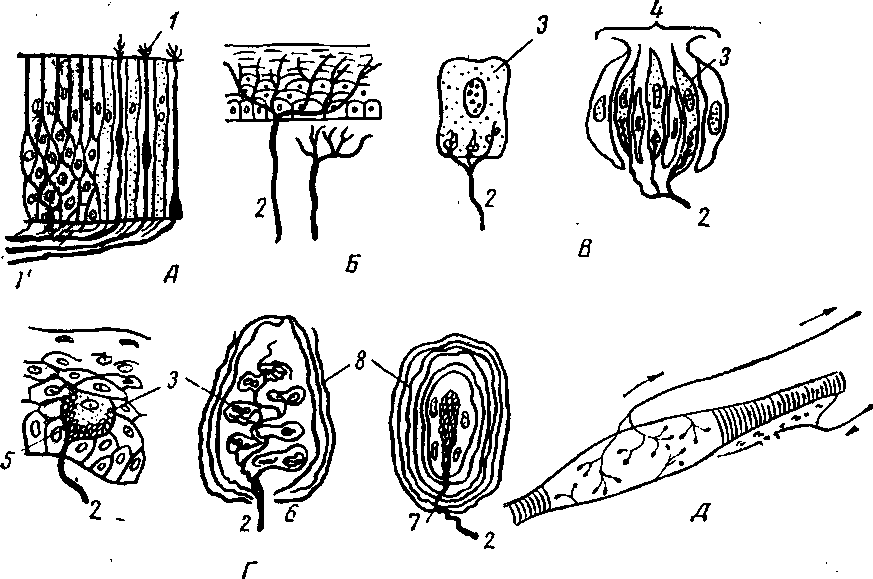
Рецепторы, несмотря на большое разнообразие в строении, могут быть свободными и несвободными нервными окончаниями (рис. 282).
Рецепторы, построенные по типу свободных нервных окончаний, представляют собой телодендрии, т. е. концевые разветвления дендритов ре-цепторных нейронов. Характер ветвления их крайне разнообразен (см. Курс гистологии). Свободные нервные окончания имеются как среди эпителиальных клеток, так и во всех других тканях. Они относятся к экстеро-и интерорецепторам.
Рецепторы, построенные по типу несвободных нервных окончаний, характеризуются разветвлением рецепторных нервных окончаний вокруг чувствительных клеток. Последние могут залегать среди эпителиальных клеток слизистой оболочки (например, клетки вкусовых луковиц языка, или неосумкованные рецепторы). Кроме того, они бывают одеты специальными оболочками, иногда очень сложной структуры — это осумкованные рецепторы (например, тельца Меркеля, Догеля, Грандри, Фатер — Пачини — для восприятия глубокого давления; Гербста, Мацони, Краузе — для восприятия температуры, тельца Мейснера и пр.). К типу осумкованных рецепторов относят также органы зрения, слуха и равновесия с очень сложным микроскопическим и макроскопическим строением.
Развитие рецепторных аппаратов анализаторов. При рассмотрении нервной системы отмечалось, что чувствительные и двигательные нервные клетки развивались из первоначально одинаковых эпителиальных клеток ' энто- и эктобласта в соответствии с «принципом разделения труда». Примитивные чувствительные клетки низших животных не обладают избирательностью, так как они одинаково реагируют на самые различные раздражители — как физические, так и химические. Под влиянием постоянно дейст-
17*
483
вующих раздражителей в процессе эволюции возникают экстерорецепторы своеобразного строения с избирательной функцией. Так, одни рецепторы воспринимают раздражения световой энергии, другие — звуковых волн, третьи — химической энергии, четвертые — различные механические раздражения. Одновременно появляются интерорецепторы, реагирующие на специфические раздражения, возникающие в различных органах и тканях, т. е. во внутренней среде организма.
У низших животных первичные чувствительные клетки широко распространены среди эпителиальных клеток, в то время как у высших животных они представлены обонятельными клетками и нейроэпителием сетчатки глаза. С превращением первичных чувствительных клеток в нервные ре-цепторная функция их сохраняется за дендритами, которые как концевые, или свободные, нервные окончания (телодендрии) разветвляются среди эпителиальных клеток кожного покрова или под ними. Такие свободные нервные окончания встречаются в большом количестве как у беспозвоночных, так и у позвоночных, причем у последних не только в кожном, покрове, но и во всех внутренних органах и тканях.
С развитием из тех же эпителиальных клеток вторичных чувствительных клеток концевые нервные окончания вступают с ними в тесный контакт. Таким образом возникают уже несвободные нервные окончания. Вторичные чувствительные клетки имеются у некоторых беспозвоночных (черви и членистоногие), однако закономерно они присущи только позвоночным животным вследствие их сложных взаимоотношений с внешней средой.
У позвоночных специальные чувствительные клетки, как и вся соматическая нервная система, происходят из общего зачатка нервной системы.