

2.2. Связь поведения с некоторыми биохимическими показателями.
В основе всех поведенческих изменений лежат нейрохимические и сопутствующие им нейрофизиологические сдвиги определенных функциональных единиц центральной нервной системы. В исследованиях по функциональной нейрохимии показано, что при любом физиологически активном состоянии нервных клеток, в том числе и в процессе обучения усиливается метаболизм РНК. Тесно связан с интенсивностью обмена РНК белковый обмен нервных клеток.
Интенсификация синтеза РНК и специфических белков в процессе функционирования нейронов дает основание полагать, что генетически детерминированные различия в поведении могут быть обусловлены особенностями метаболизма РНК и белков в определенных функциональных отделах мозга. Например, были отмечены межлинейные различия по содержанию РНК и некоторых участвующих в ее метаболизме ферментов у линий крыс, различающихся по скорости образования рефлексов. Выявлены также межлинейные особенности обмена РНК у крыс, предрасположенных к аудиогенным эпилептическим припадкам (линия Крушинского — Молодкиной), и не предрасположенных. У первых количество РНК на нейрон ядра Дейтерса достоверно выше, чем у вторых. Причем эти различия сказываются еще до появления в ходе онтогенеза патологической реакции на звук, т. е. они не являются следствием этой реакции, а, вероятно, генетически обусловлены.
Генетические различия в скорости условнорефлекторного обучения могут быть до некоторой степени скоррелированы с интенсивностью синтеза специфичных для всей нервной ткани белков.
То, что поведенческая изменчивость обусловлена некоторыми метаболическими эффектами генов внутри нервной клетки, подтверждают также данные по связи между ферментами и поведением. Так, обнаружены линейные различия у мышей по активности фенилаланингидроксилазы, которая в норме превращает фенилаланин в тирозин, и скоррелированные различия в поведении. Активность этого фермента у гомозигот по гену дайльют (dilutedd), характеризующихся ослабленной пигментацией и крайней нервозностью, уменьшена почти наполовину. Летальные же гомозиготы по гену дайльют (d1d1) проявляют только 14% нормальной активности фенилаланингидроксилазы. Названные мутации подавляют активность фенилаланингидроксилазы через продукцию ингибитора фермента, который у нормальных мышей уменьшается на третьей постнатальной неделе.
Биохимия алкоголизма также демонстрирует корреляцию между активностью ферментной системы, и поведением. Оказывается, что различные линии мышей в условиях предоставления им выбора воды и алкогольных растворов различной концентрации, характеризуются стабильными линейными средними употребленного алкоголя. Выявлена корреляция между этими средними и активностью печеночной алкогольной дегидрогиназы (АДН). Линия с сильным алкогольным предпочтением характеризовалась высоким уровнем активности АДН. И наоборот, линия с сильным избеганием алкогольных растворов имела низкую активность фермента.
В условиях различных алкогольных режимов у животных всех линий наблюдалось адаптивное изменение активности фермента, но характер этого изменения был специфичным для каждой линии.
В экспериментах показано, что моноаминэргическая система мозга принимает участие в регуляции многих форм поведения у человека и животных в норме и патологии. Исследователи предполагают, что уровень экспрессии генов тирозингидроксилазы (TH), дофаминового транспортера (DAT), моноаминоксидазы А (MAO-A) и серотонинового транспортера (SERT) может играть определенную роль в таких поведенческих процессах, как агонистическое (конкурентное) и субмиссивное (подчиненное) поведение.
Методом полуколичественной мультиплексной полимеразной цепной реакции ученые исследовали изменение экспрессии генов SERT и МАОА в ядрах шва среднего мозга, генов TH и DAT в VTA мозга мышей линии СВА/Lac c агрессивным и подчиненным типами поведения. По результатам двух независимых экспериментов у подчиненных животных обнаружено значительное увеличение уровня мРНК генов МАОА и СЕРТ по сравнению с контрольной группой и агрессивными особями. Вероятно, увеличение экспрессии генов МАОА и СЕРТ отражает ускоренную инактивацию серотонина в ответ на усиление функционирования серотонергической системы под влиянием социального стресса.
В тоже время у мышей линии СВА/Lac c агрессивным типом поведения было обнаружено достоверное увеличение уровня мРНК генов TH и DAT в мезолимбических дофаминэргических нейронах мозга.
Эксперименты показали, что хроническое проявление агрессии сопровождается долговременной активацией дофаминэргической системы, в частности активацией генов TH и DAT, вовлеченных в синтез и инактивацию дофамина, соответственно.
Ген, кодирующий фермент моноаминоксидаза А (MAOA), который разрушает нейротрансмиттеры, оставшиеся в синаптической щели, привлек внимание ученых, когда была обнаружен семья с мутацией в этом гене.
История такова. На прием к врачу пришла женщина, которая хотела иметь ребенка, но боялась, что ребенок будет болен, поскольку у нее в семье мужчины (не все, но многие) отличались высокой агрессивностью (немотивированная агрессия и легкая степень умственной отсталости). Когда они были обследованы, оказалось, что это не просто дурной характер, а генетически детерминированное заболевание. Родословная этой семьи представлена на рисунке (кружочки с ромбиком в центре обозначают женщин-носительниц мутации).
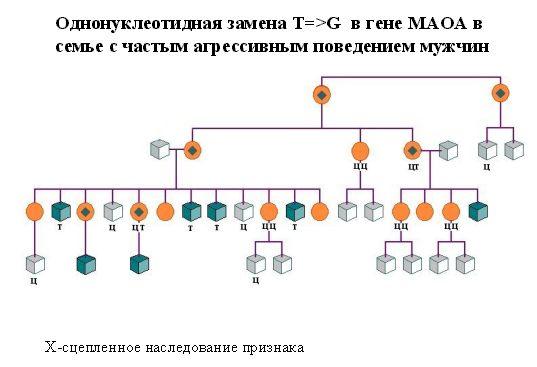
Рис. 2.5. Однонуклеотидная замена Т—Г в гене МАОА в семье с частым агрессивным поведением мужчин
По характеру наследования поняли, что это заболевание, сцепленное с полом, потому что здоровые женщины рожали больных сыновей (дочери рождались нормальными, то есть, не были агрессивными).
В Х-хромосоме обнаружили мутантный ген, и выяснили, что он кодирует МАОА. Мутация – единичная нуклеотидная замена, которая приводила к обрыву синтеза белка и МАОА при этом не функционировала.
Таким образом, замена одного нуклеотида (цитозина на тимин в данном случае) приводила к ярко выраженному аизменению поведения.
Этот ген стали более активно исследовать. Такая мутация, как в этой семье, полностью инактивирующая фермент, больше нигде не была найдена, но были обнаружены мутации, снижающие активность МАОА.
Ученые исследовали группу людей с низкой и нормальной активностью МАОА и сравнили количество случаев асоциального поведения (нападение, нападение с целью нанесения повреждений, убийство, изнасилование, жестокость по отношению к животным). Когда просто так исследовали выборку, разницы не обнаружили. Но когда эту выборку разделили по условиям воспитания в детстве (Рис. 2.6), то оказалось, что разница в поведении людей, которые воспитывались в благоприятных условиях, практически отсутствует, а вот если условия были плохие, то разница становится существенной.
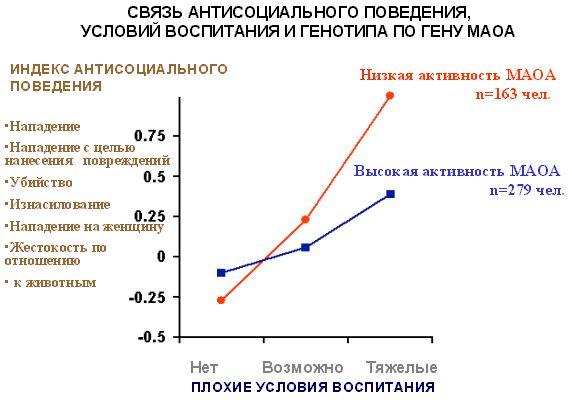
Рис. 2.6. Связь асоциального поведения, условий воспитания и генотипа по гену МАОА.
Для изучения связи моноаминоксидазы А с особенностями дповедения провели серию экспериментов на мышах. У мышей «нокаутировали» ген МАОА. Такие мыши были очень агрессивны, они набрасывались на своих сородичей без всякого повода, то есть вели себя также, как и люди с аналогичным генетическим дефектом.
Подобные исследования помогают понять, что именно в поведении детерминировано генетически и в какой мере, а что подвержено влиянию среды.
Все биохимические различия у животных, характеризующихся разным поведением, могут обусловливаться не только истинными генетическими корреляциями, но и целым рядом сложных взаимодействий, как в цепях метаболизма, так и со средой.
Но большинство наблюдаемых наследственных различий в специфическом поведении не ассоциируется со специфическими белковыми молекулами или ферментами. Поведенческая изменчивость детерминируется в основном количественной регуляцией функционального состояния тех или иных отделов центральной нервной системы.
Множество факторов могут регулировать функциональное состояние конкретных нейронных структур, множество биохимических звеньев лежит в основе их функционирования, но, вероятно, все они выходят на уровень мембранных процессов, генерирующих нервный импульс. Генетическая изменчивость этих процессов, протекающих в специфических нейронных цепях, может детерминировать изменчивость многих форм поведения.