

5. 3. Генетика поведения насекомых
5.3.1. Насекомые как объект генетики поведения.
Для генетического анализа наиболее удобны те поведенческие реакции, развитие которых защищено от средовых колебаний. В этом отношении насекомые являются хорошим объектом для изучения генетики поведения.
В классе насекомых функции нервной системы достигают максимально возможного для беспозвоночных развития. Это относится главным образом к наследственно фиксированным реакциям. Однако некоторые из насекомых обладают и индивидуальной пластичностью поведения, т. е. способностью к образованию условных навыков. Особенно это характерно для представителей отряда перепончатокрылых (Hymenoptera), поскольку они имеют сложную организацию общественной жизни.
В нервной системе насекомых, так же как и у позвоночных, различают три типа нейронов: чувствительные, двигательные и ассоциативные. Чувствительные нейроны, как правило, локализованы в органах чувств, двигательные входят в состав нервных центров, а их аксоны заканчиваются в том или ином эффекторе. Ассоциативные нейроны, локализованные в грибовидных телах, осуществляют передачу возбуждения с нейрона на нейрон. Эта передача имеет электрохимическую природу и осуществляется в области синапсов с помощью химических медиаторов.
Насекомые имеют хорошо развитые органы чувств.
Орган зрения состоит обычно из фасеток, число которых варьирует от нескольких единиц до тысяч. Этот орган гораздо лучше приспособлен к восприятию движения, чем к восприятию формы. Кроме того, многие насекомые могут воспринимать ультрафиолетовое излучение как особый цвет.
Орган слуха очень разнообразен по строению. Иногда он представляет собой щетинку, в основании которой находится чувствительная клетка, иногда эта чувствительная клетка располагается на голенях передних ног, в местах утонченного в виде барабанной перепонки участка кутикулы.
Особенность органа слуха насекомых заключается в том, что он может воспринимать колебания вообще (например, резонансные колебания почвы), а также ультразвуки.
Хорошо развиты у насекомых хеморецепторы, которые чаще всего представляют собой простые щетинки, улавливающие вкусовые и обонятельные раздражения.
Насекомые, особенно перепончатокрылые, проявляют очень интересные формы поведения, например при ориентации, при строительстве гнезд и т. д. Наивысшего развития поведение достигает у общественных насекомых, наиболее изучено оно у пчел.
Все это, а также короткий жизненный цикл насекомых и изученность частной генетики некоторых из них (например, дрозофилы) делают их интересным и удобным объектом для генетического анализа поведения.
5.3.2. Влияние отдельных генов на поведение насекомых
Наследственно фиксированные, или инстинктивные, реакции насекомых, состоящие из отдельных двигательных актов, являются фенотипами для генетического анализа. Ученые исследуют как эффект отдельных мутаций, так и результат селекции на выполнение этих реакций.
Признаки, находящиеся под контролем единичных генов, изучены лучше других, потому что их проще всего наблюдать. Это относится к любым признакам, будь то морфологические, физиологические или поведенческие свойства организма. К таким генам относятся, прежде всего, гены, изменяющие и внешний вид животных и одновременно их поведение, то есть, гены, имеющие плейотропное действие.
Классическим примером поведения, определяемого единичным геном, является гигиеническое поведение пчел.
Ротенбулер(Dr. Walter C. Rothenbuhler) провел в 1964 году изящный анализ процесса очистки пчелами сот от личинок, погибших от распространенной болезни — американской пчелиной гнильцы (переносчик— Bacillus larvae). Он обнаружил пример поведения, имеющего явное приспособительное значение и контролируемого двумя единичными локусами.

Рис.5.10. Ротенбулер (Dr. Walter C. Rothenbuhler)
Поддержание гигиенических условий в улье требует распечатки ячеек, в которых находятся пораженные болезнью личинки, и немедленного их изъятия. В противном случае трупы личинок с содержащимися в них спорами переносчика болезни остаются в улье и служат постоянным источником распространения инфекции.

Рис.5.11. Соты, пораженные американской пчелиной гнильцой
Гигиеническое (или не гигиеническое) поведение пчел определяется генами, расположенными в двух независимо расщепляющихся локусах. Один из генов ответствен за поведение, связанное с распечаткой сотовых ячеек, другой — за поведение, связанное с удалением их содержимого. Если обозначить буквой u рецессивный ген, управляющий распечаткой ячеек, а буквой r — рецессивный ген, ответственный за изъятие их содержимого, то генотип пчел в гигиеническом улье имеет вид uurr, а генотипы пчел, не выполняющих гигиенических действий - UURR, UuRr, UURr, UuRR.
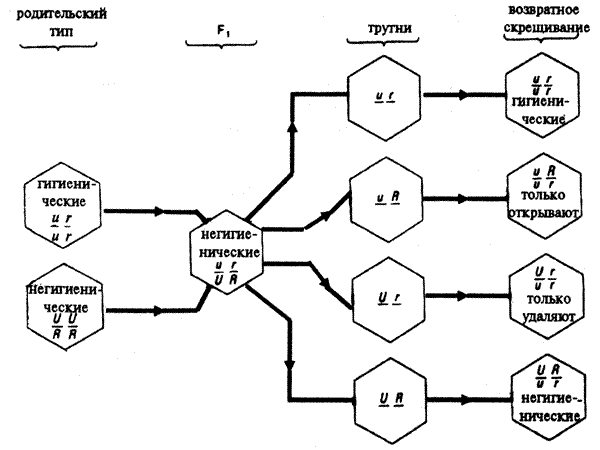
Рис.5.12. Схема наследования признака «гигиеническое поведение» у медоносной пчелы.
Движения пчел, которые они совершают при вскрытии ячеек и выбрасывании зараженных личинок, относятся к категории ФКД и обладают всеми характерными для них признаками.
Данные движения выполняются насекомыми в полной форме без предварительного опыта и практически неизвестно, чтобы они видоизменялись при «тренировке». Более того, эти движения неспецифичны и характерны для вида в целом, а не только для каких-либо пород пчел. Все пчелы выполняют эти движения, однако у пород, подверженных заболеванию, частота этих ФКД чрезвычайно низка. Поэтому их поведение не может играть своей адаптивной роли: зараженные личинки не удаляются, и инфекция распространяется
Эксперименты показали, что на гигиеническое поведение рабочих пчел двух инбредных линий - устойчивой к американскому гнильцу и восприимчивой к нему - не влияют такие факторы, как возраст, число погибших от инфекции личинок и характер питания. Первое поколение от скрещивания этих двух линий оказалось восприимчивым к американскому гнильцу, т. е. гибридные рабочие пчелы не очищают улей от больных личинок.
В результате анализирующего скрещивания гибридных особей с устойчивой родительской линией было получено четыре одинаковых по численности типа рабочих пчел:
полностью проявляющие гигиеническое поведение, т. е. вскрывающие ячейки с больными личинками и выбрасывающие их из улья;
совсем не проявляющие этих форм поведения;
вскрывающие ячейки с больными личинками, но не выбрасывающие их;
не способные вскрывать ячейки с больными личинками, но выбрасывающие эти личинки после того как экспериментатор вскроет ячейки.
Эти данные подтверждают дигибридный характер наследования гигиенического поведения пчел. Напомним, что ген u в гомозиготном состоянии детерминирует способность вскрывать ячейки с больными личинками, а ген U – определяет неспособность это делать. Ген r в гомозиготном состоянии контролирует способность выбрасывать больные личинки из вскрытых ячеек, а ген R - определяет неспособность это делать. Тогда становятся понятными генотипы особей, со всеми вариантами поведения.
Генотип пчел, полностью проявляющих гигиеническое поведение, т. е. вскрывающие ячейки с больными личинками и выбрасывающие их из улья - uurr. Генотипы пчел, совсем не проявляющих этих форм поведения - UURR, и другие генотипы, содержащие оба доминантных гена. Генотипы пчел вскрывающих ячейки с больными личинками, но не выбрасывающие их - uuRR, uuRr. Генотип пчел не способных вскрывать ячейки с больными личинками, но выбрасывающие эти личинки, после того как экспериментатор вскроет ячейки - UUrr, Uurr.
Был выявлен ряд генов, уровень экспрессии которых меняется у рабочих пчел в зависимости от рода их занятий. Особо интересен среди них ген vitellogenin. Этот ген у насекомых — как общественных, так и одиночных — тесно связан с женской репродуктивной функцией: кодируемый им белок вителлогенин необходим для формирования яиц в яичниках самок. Однако у рабочих пчел, которые никаких яиц не откладывают, этот ген тоже активен. Более того, было замечено, что его активность у рабочих пчел снижается с возрастом. Это позволило предположить, что вителлогенин служит своеобразным поведенческим «переключателем»: снижение его концентрации заставляет пчелу в определенный момент ее жизни оставить домашние хлопоты и переключиться на сбор пропитания за пределами гнезда.
Группе американских энтомологов удалось экспериментально подтвердить роль вителлогенина в регуляции разделения труда у пчел. Для этого они воспользовались методом РНК-интерференции. Только что вышедшим из куколки молодым рабочим пчелам вводили двухцепочечную РНК с такой же последовательностью нуклеотидов, как и в гене vitellogenin. В результате этот ген у подопытных пчел оказывался выключенным на всю жизнь. В опыте использовались также две контрольные группы рабочих пчел: первой из них ничего не вводили, а второй вводили другую двухцепочечную РНК, не влияющую на экспрессию генов (чтобы доказать, что наблюдаемые изменения вызваны именно выключением гена vitellogenin, а не травмирующей процедурой).
Оказалось, что рабочие пчелы с отключенным геном vitellogenin отличаются от контрольных по трем показателям.
Во-первых, они значительно раньше переходили от домашних работ к сбору пропитания. Это подтвердило высказанное ранее предположение, что вителлогенин служит «поведенческим переключателем».
Во-вторых, продолжительность их жизни заметно сократилась.
В-третьих, они специализировались преимущественно на сборе нектара, а не пыльцы.
Таким образом, один и тот же ген, изначально связанный с женской репродуктивной функцией, стал выполнять у медоносной пчелы целый ряд новых функций, связанных с регуляцией разделения труда и общественного устройства.
Проявляется влияние отдельных генов на другие аспекты поведения насекомых. Мутантные гены cinnabar и vermilion не только изменяют цвет глаз Drosophiia melanogaster, но влияют также на скорость спаривания. В экспериментах самцы vermilion спаривались менее успешно, чем самцы cinnabar. Мутация yellow изменяет не только цвет тела, но и брачное поведение самцов.
Гомозиготы D. melanogaster по генам v/bw (vermilion/brown) имеют светло-вишневые глаза и сцепленную с этим признаком заметно сниженную остроту зрения, что проявляется в слабой двигательной активности в ответ на световые раздражители. В популяции, содержащей мух дикого типа и мух с геном Ваг, самцы Ваг уступают самцам дикого типа в успехе при спаривании.
Есть и другие формы поведения D. Melanogaster, контролируемые единичными генами. Это мутации затрагивающие реакцию на стрессовое воздействие, нейромышечное поведение, локомоцию и зрение, циркадные ритмы.
Например, Сеймур Бензер с сотрудниками изучили три мутации единичных генов, изменяющих нормальный (примерно 24-часовой) циркадный ритм активности, характерный для D. Melanogaster. Мухи этого вида обычно вылупляются на рассвете, когда выпадает роса и влажность воздуха повышается, что отвечает свойственной дрозофиле потребности во влажных условиях среды. Отсюда и название Drosophila, что значит по латыни — «любящая росу». После вылупления у большинства видов дрозофилы наступает период утренней активности, который продолжается до полудня, а вечером наблюдается еще одна короткая вспышка активности.

Рис.5.13. Сеймур Бензер
Убедительным свидетельством в пользу того, что действие биологических часов находится под генетическим контролем, служит существование вылупляющихся в любое время суток аритмических мутантов с укороченным периодом, для которых характерен не 24-часовой, а 19-часовой ритм, и мутантов с удлиненным периодом, обладающих 28-часовым ритмом.