

6.7. Использование мозаиков для выявления структур, затронутых поведенческими мутациями.
Известно, что мозаичные особи, имеющие генотипически различные части тела, представляют огромную ценность для исследования клеточной автономии, специфичности выражения генов в различных тканях, взаимодействия тканей разного генотипа и др.
В плане генетики поведения систематическое изучение мозаиков D. melanogaster началось по инициативе С. Бензера. В 1970 г. ХХ века он вместе с И. Хота опубликовал статью под названием «Генетическое рассечение нервной системы дрозофилы посредством мозаиков». В ней описано несколько способов получения мозаиков, из них наиболее эффективен метод утраты в онтогенезе дрозофилы кольцевой Х-хромосомы.
В качестве исходного материала брали гетерозиготных особей на стадии зиготы, у которых одна Х-хромосома кольцевая — In(l)wvC, а другая, анализируемая, несет выбранную поведенческую мутацию и цепочку рецессивных маркеров у w sn f. Можно взять и другие маркеры, затрагивающие покровы, окраску тела и глаз, форму щетинок.
Вероятность разрыва и утраты кольцевой Х-хромосомы In(l)wvC в процессе клеточных делений очень велика и составляет от 10% до 50%. В результате этого и образуются мозаичные особи, которые содержат исходные клетки дипло-Х и возникшие в результате потери кольцевой Х-хромосомы клетки гапло-Х с единственной Х-хромосомой (анализируемой).
Чем раньше в онтогенезе произойдет разрыв и утрата Х-хромосомы In(l)wrC, тем большее количество клеток тела мухи будет гапло-Х. Если это событие произойдет уже при первом дроблении зиготы, то образуется гинандроморф, у которого одна половина тела будет женской, с клетками дипло-Х, а другая — мужской, с клетками гапло-Х. Участки тела с клетками гапло-Х легко узнаются по рецессивным маркерам, проявляющимся в гемизиготном состоянии.
Дробление у дрозофилы происходит по синцитиальному типу, и примерно после 12 делений ядра мигрируют к поверхности яйца, образуя бластодерму. На стадии бластодермы отдельные ее участки уже детерминированы. Из них формируются имагинальные диски в теле личинки, которые, в свою очередь, дают начало вполне определенным частям тела имаго. Отдельные участки поверхности тела (экзоскелета) и прилежащие к ним ткани с высокой вероятностью состоят из клеток одного и того же клона. Например, если левая часть переднего грудного отдела мухи имеет кутикулу желтого цвета, т. е. состоит из клеток гапло-Х, то имеются все основания полагать, что соответствующий отдел центральной нервной системы — левая часть грудного синганглия — также будет с высокой вероятностью образована клетками гапло-Х. Это позволяет находить так называемые фокусы — структуры, затрагиваемые анализируемыми мутациями.
В работе, посвященной мутациям дефектов зрения, Хотта и Бензер пришли к важному выводу об автономности зрительной системы каждого глаза. При снятии ЭРГ у билатеральных гинандроморфов рецепторный потенциал нормального глаза оставался нормальным, а рецепторный потенциал мутантного глаза был аномальным. Удалось отобрать таких мозаиков, все тело которых было нормальным и только один глаз являлся мутантным, т. е. состоял из клеток гапло-Х с анализируемой мутацией в Х-хромосоме. ЭРГ, снимаемая с такого глаза, неизменно оказывалась дефектной, характерной для данной мутации.
К. Икеда и В. Каплан, пользуясь аналогичной методикой, исследовали проявление шейкер-мутации HklP у 77 мозаичных особей. При этом с помощью микроэлектродной техники характеризовали электрическую активность отдельных мотонейронов. Ими убедительно доказана автономность отдельных нейронов торакального синганглия. Без получения мозаичных особей подобного принципиального результата вряд ли удалось бы достичь.
Хотта и Бензер в разных экспериментах анализировали начальные этапы ухаживания (преследование самок и вибрацию крыльев) и конечные этапы — попытку спаривания и само спаривание. Всего ими изучено в первой части работы 477 и во второй 208 мозаичных особей с самым различным сочетанием клеток дипло-Х и гапло-Х.
В первой части работы наблюдения вели с помощью бинокулярной лупы над отдельными парами особей гинандроморфов и самкой дикого типа C-S, которых помещали под часовое стекло. Результаты исследований показали, что контроль за ориентацией мозаиков по отношению к самкам и вибрацией крыльев в качестве элемента ухаживания осуществляется церебральным ганглием. Мозаичные особи, у которых все тело, за исключением головы, было женским, тем не менее, вели себя как самцы — преследовали самок и вибрировали крыльями.
Во второй части работ наблюдения за спариваниями проводили в приборе, включающем сразу 10 камер. Ученые обнаружили, что попытки к спариванию совершают гинандроморфы, у которых и голова, и торакс — самцовые. Для спаривания требуется к тому же нормальное мужское строение гениталий.
В дополнительной серии экспериментов к мозаичным особям подсаживали самцов дикого типа. Наличие у мозаиков женских брюшка и гениталий провоцировало ухаживание самцов. Однако чтобы мозаичная особь вела себя как самка, ее центральная нервная система должна была оставаться женской.
Характеристики мозаиков.
Мозаики — это сложные особи, у которых некоторые участки различных тканей могут быть мутантными, в то время как остальные имеют нормальный генотип. Это дает возможность идентифицировать ту часть, которая служит «точкой приложения» мутантного гена при формировании мутантного поведенческого фенотипа особи.
Половые мозаики (или гинандроморфы), у которых одни части тела имеют генотип самца, а другие — самки, могут дать интересную информацию относительно организации полового поведения.
Первые работы в этом направлении относятся еще к 1932 году, когда Уайтинг изучал половое поведение паразитических ос (Habrobracon). У них различие в половом поведений не сводится, как у дрозофилы, к отсутствию у самок реакций, характерных для самцов. Наблюдая гинандроморфов, которые имеют голову одного пола, а остальную часть тела — другого, Уайтинг обнаружил, что поведение определяется генотипом клеток головного ганглия. Билатеральные (лево-правые) мозаики проявляли тенденцию как к поведению, характерному для самцов, так и к поведению, характерному для самок.
Имеется несколько генетических ситуаций, ведущих к образованию гинандроморфов, а, следовательно, и несколько методов получения мозаиков. Один из них основан на использовании линии дрозофилы с кольцевой X-хромосомой и зависит от потери неустойчивой кольцевой X-хромосомы во время первого деления ядра зиготы эмбриона самки. В этом случае возникает два клона клеток: ХО и XX (рис. 6.3.)
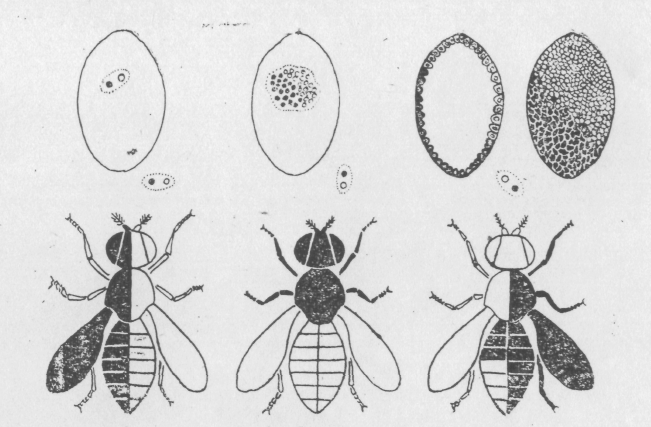
Рис.6.3. Соматическая диаграмма образования генетических мозаиков.
Соотношение частей организма, имеющих генотипы XX и ХО, зависит от того, когда утрачивается кольцевая X хромосома. Если, например, она утрачивается при первом делении ядра зиготы, то клеточные клоны занимают по половине бластодермы, если позднее, то клон ХО занимает меньшую часть бластодермы. В редких случаях кольцевая X-хромосома утрачивается при первом делении и еще раз при более поздних делениях, в этих случаях у эмбрионов клон ХО занимает большую часть бластодермы.
Характер же мозаицизма будет определяться не только осью первого деления, но и характером миграции ядер к поверхности при образовании бластулы. Относительное положение ядер имеет тенденцию сохраняться, но все-таки при миграции ядра с генотипом ХО могут мигрировать вместе с ядрами с генотипом XX.
Мигрирующие ядра равнонаследственны, и при образовании бластодермы место, занимаемое клеткой, в основном определяет ее судьбу, т. е. те структуры, происхождение которым она даст. Этот механизм обеспечивает появление самых различных мозаиков.
Нервная система и мезодерма, дающая образование мышечной ткани, также образуются из специфических районов бластодермы. Границы у мозаиков между «мужскими» и «женскими» частями в основном совпадают с межсегментальными границами и продольной средней линией экзоскелета (рис. 6.2.), так как экзоскелет есть совокупность многих частей, образующихся независимо в ходе метаморфоза из имагинальных дисков личинки. Имагинальные диски, в свою очередь, образуются из специфических районов бластодермы. Однако отдельные клеточные группы внутри одного диска могут быть также разного генотипа.
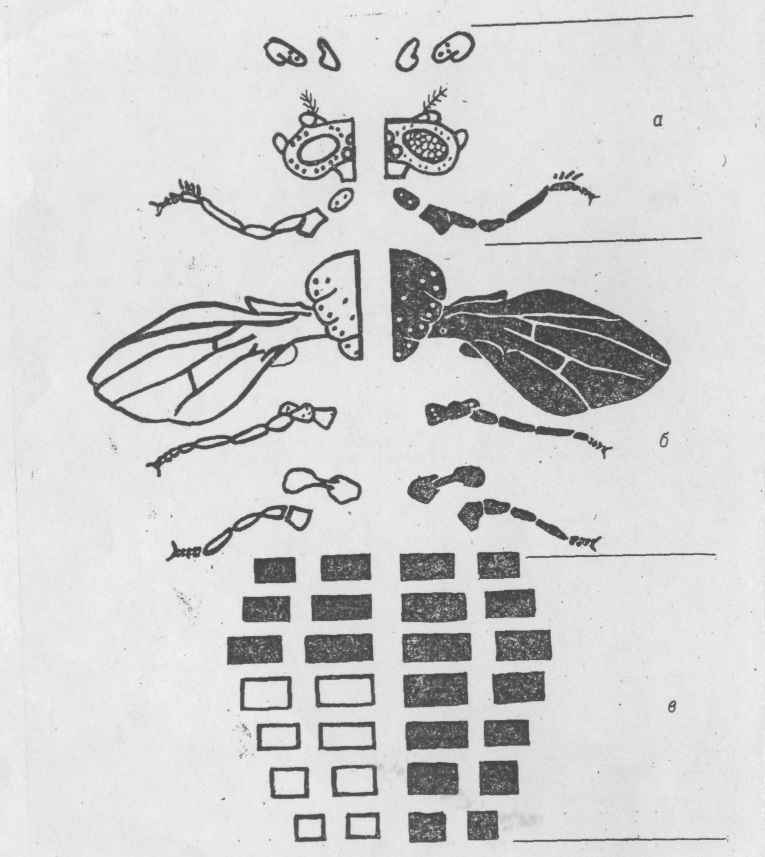
Рис. 6.4. Схематическое изображение внешних частей тела дрозофилы, образующихся независимо из отдельных имагинальных дисков.
а —голова; б —грудь; а — брюшко.
Для маркирования «мужских» и «женских» частей тела используются генетически-рецессивные анатомические маркеры (желтый цвет тела, белые глаза, вильчатые щетинки и т. д.). Маркеры не проявляются в тех частях тела, которые имеют генотип XX. В этом случае они прикрыты нормальными аллелями. Однако они видны в «мужских» частях тела, утративших кольцевую X-хромосому и имеющих одну X-хромосому. Благодаря этому, отбирая различных мозаиков, можно найти, какие части тела должны быть мутантными, чтобы сформировался мутантный поведенческий фенотип. Иными словами, можно установить, какая именно анатомическая структура или клеточная ассоциация должна быть затронута мутацией, чтобы проявился эффект мутации на уровне целостного организма.
Разрешающая способность этого метода зависит от числа исследуемых особей: чем больше это число, тем больше вероятность, что среди них будут мозаики по самым ближним точкам.
Используя мозаиков, С. Бензер исследовал самые разнообразные мутации, например серию нефототаксисных мутаций, которые принадлежали пяти различным цистронам X-хромосомы. В норме мухи характеризуются положительным фототаксисом и двигаются по направлению к свету прямо, при этом свет равной интенсивности падает на оба глаза. Мутантные мухи с отрицательным фототаксисом предпочитают двигаться не к свету, а от него. Нефототаксисные же мутанты не проявляют ни положительного, ни отрицательного фототаксиса.
Интересно поведение билатеральных мозаиков у которых один глаз дефектный, т. е. мутантный, а другой — нормальный. Они реагируют на свет, но в отличие от нормальных мух летят по направлению к источнику света не прямо, а по спирали, постоянно поворачивая свой мутантный глаз к свету, чтобы интенсивность света, падающего на оба глаза, была одинаковой (рис. 6.5.).
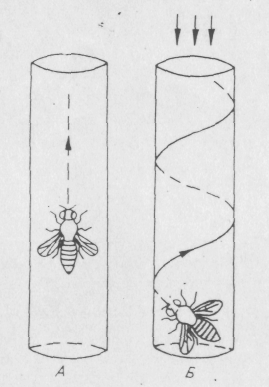
Рис. 6.5. Движение к источнику света билатеральных мозаиков по фототаксисным мутациям.
А — нормальная муха; Б — билатеральный мозаик с одним мутантным глазом.
Исследование электроретинограммы мутантных по поведению мух подтвердило, что первичный фокус действия большинства нефототаксисных мутаций находится в самом глазу. Все мутации вызывали различные отклонения в электроретинограмме, так как мутантный эффект у разных мутаций осуществлялся разными путями. Причина одних отклонений — пониженная светочувствительность фоторецепторных клеток, другие мутации вызывали дегенерацию зрительных палочек, у третьих — отклонения в электроретинограмме обусловливались дефектом в передаче нервного сигнала от рецепторных клеток следующим нейронам.