

Петров_К_М_Биогеография_СПб_2001
.pdfРис. 3.7. Личиночные стадии морских беспозвоночных, проходящих планктонную стадию развития, по Ж. П. Эрхарду и Ж. Сежену:
а – личиночные стадии морского ежа; б – личиночные стадии многощетинкового червя; в – личиночные стадии краба
Это заставляет их совершать дальние перелеты (рис. 3.8). Также не совпадают районы размножения и питания многих морских рыб, пресмыкающихся (рис. 3.9). Дальние миграции совершают киты; регулярные странствования осуществляют ластоногие от районов нагула на строго определенные места размножения.
Непреднамеренное распространение животных человеком подчас приносит большой вред. Особенно это касается разного рода вредителей и паразитов. Спутниками человека являются домовые мыши, крысы, тараканы и пр. Колорадский жук, завезенный из Америки, вредит посадкам картофеля в Европе. Европейский кролик, попавший в Австралию в 1840 г. в количестве 16 особей, вскоре размножился там настолько, что превратился в настоящее бедствие. Он стал серьезным конкурентом на пастбищах как местным травоядным, так и разводимым фермерами овцам.
3.2. Географические и генетические элементы биоты
Виды организмов, совокупность которых образует биоту, различны в отношении
их географического распространения и происхождения. Поэтому при характеристике биоты важным аспектом исследования является изучение закономерностей распространения видов и определение мест их вероятного происхождения.
В соответствии с особенностями распространения видов выделяются географические элементы биоты. Ареалы географических элементов в основном формируются в пределах зональных типов ландшафтов или физико-геогра- фических регионов, обладающих единством палеогеографического развития (рис. 3.10).
Выяснение того, откуда происходят виды, как они попали в состав изучаемой биоты, предполагает выделение генетических элементов. Прекрасным примером такого анализа является заключение известного русского ботаника А. Н. Краснова о связи тундровых кус-
тарничков с тропической растительностью гор Юго-Восточной Азии: “… близость флоры горных вершин Явы к столь противоположным по свойствам тундрам севера представляет громадный интерес, так как она показывает, что флора тундры может быть непосредственным производным флоры тропического леса” (Краснов, 1987).
Любая биота состоит из многих географических элементов. Например, А. А. Гроссгейм приводит следующее соотношение элементов флоры Кавказа (в %):
Кавказские (собственно) – 20,0 Бореальные (северные) – 38,2 Средиземноморские – 13,2 Колхидские - 12,7 Переднеазиатские – 9,7 Понтические – 5,6 Туранские – 0,1 Пришлые виды – 0,5
Данный спектр элементов наглядно демонстрирует их роль в формировании флоры. Широкое участие бореальных элементов в биотах отражает внедрение северных видов на Кавказ в ледниковую эпоху. Свой
51
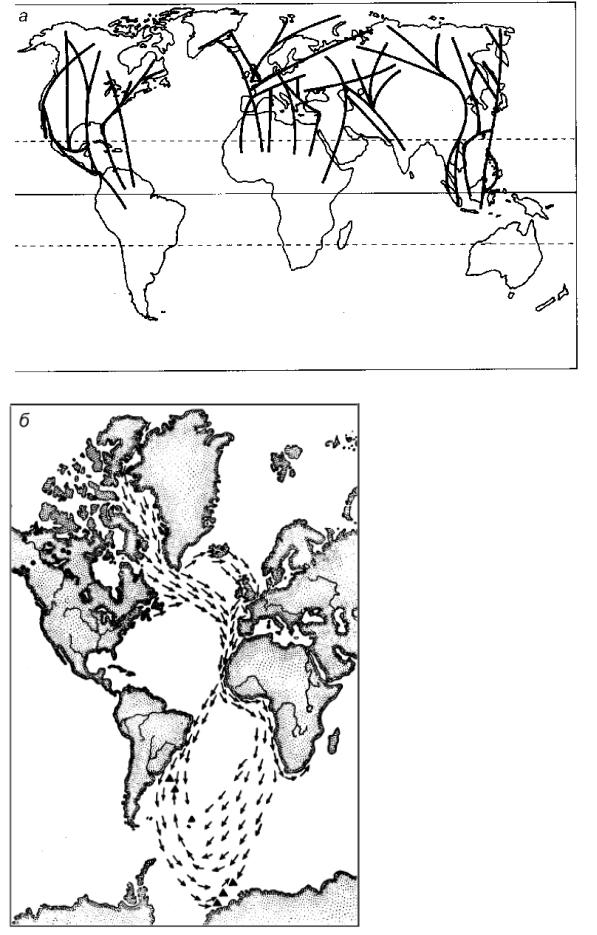
Рис. 3.8. Маршруты перелетных птиц:
а – главнейшие маршруты, по J. A. Brown a. A. C. Gibson. Они ориентированы с севера, от мест гнездования, на юг, к местам зимовок; б – распространение и перелетные пути американской популяции длиннохвостой крачки, по Н. А. Бобринскому. Черные кружочки – места гнездования, треугольники – места зимовок, стрелки – направление и пути пролета основной массы особей на зимовку
52
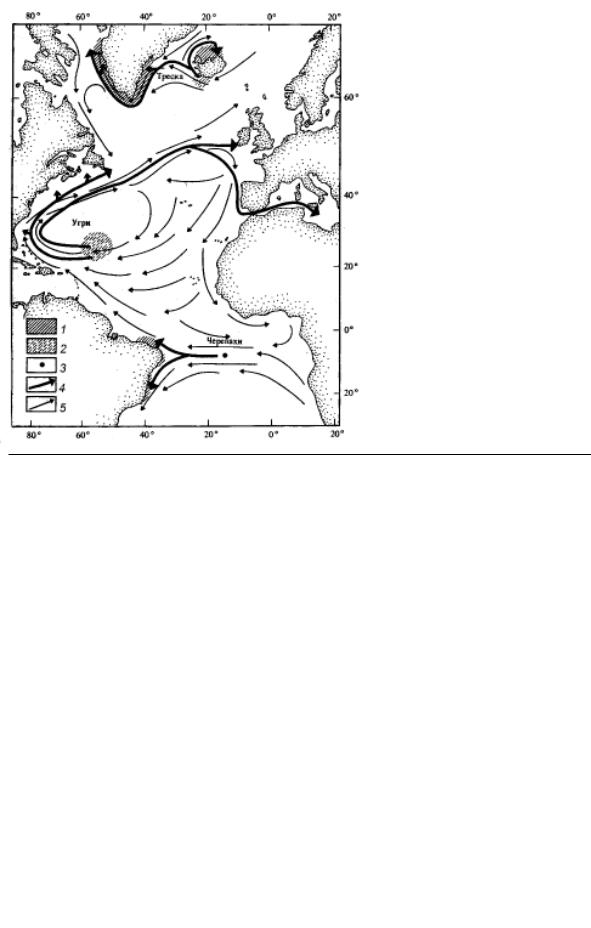
Рис. 3.9. Пути миграции трески, угрей и морских черепах, по С. Нешиба:
1 – области питания; 2 – районы нереста; 3 – район гнезд черепах; 4 – пути миграции; 5 – поверхностные океанические течения
Рис. 3.10. Европейские географические элементы флоры, по Г. Вальтеру:
а – схема основных географи- ческих элементов флоры Средней Европы; б – распространение важнейших географических элементов флоры Европы. Обозначения элементов те же, что на схеме: sm - субсредиземноморский элемент; submi – среднеевропейский + среднерусский элементы; arct – арктический; bor (w) – бореальный западный; bor (o) – бореальный восточный; atl (n) – атлантический северный; atl (s) – атлантический южный; ро – понтический; tur – туранский; medit (w) – западносредиземноморский; medit (o) – восточно-средиземноморский. Горы не приняты во внимание. Без обозначений – две сухие области: паннонская в Венгрии и бассейн Эбро в северовосточной Испании (небольшая сухая область в юго-восточной Испании не очерчена)
53
вклад в обогащение флоры внесли соседние регионы. Однако весьма значительна доля собственно кавказских видов, свидетельствующая о том, что Кавказ представляет собой самостоятельный центр видообразования.
3.3. Роль естественных барьеров
Географическое распространение видов тесно связано с их экологической пластичностью. По способности выдерживать колебания основных экологических факторов организмы делятся на эврибионтные и стенобионтные. Первые способны жить в условиях широкой амплитуды экологических факторов, вторые выносят их изменения лишь в узком диапазоне. Соответственно эврибионтные виды способны стать эврихорными, сформировать широкий географический ареал. Стенобионтные виды, являются стенохорными и имеют узкие ареалы.
Заметим, что эврихорность свойственна только немногим видам-эврибионтам. Сообщества организмов всегда более или менее стенотохорны. Наиболее широким ареалом обладают зональные типы сообществ: дождевые тропические леса, саванны, степи, тайга и т. д. В виде островков определенные сообщества могут заходить в соседние зоны (явление экстразональности), но никогда вне своей зоны они не будут доминирующими: байрачные дубовые леса в степной зоне, участки широколиственных лесов в южной тайге, острова лугов в широколиственных лесах.
На пути неограниченного увеличения численности популяций, давления жизни и стремления расширить ареал встают внешние факторы: географические, ландша- фтно-экологические и биологические.
В качестве географических факторов
выступают крупные элементы строения земной поверхности, играющие роль преград на пути расселения организмов. Для сухопутных растений и животных такими преградами являются горные хребты, океаны и моря; для водных - обширные пространства суши.
Некоторые физико-химические параметры внешней среды, играя роль экологи- ческих факторов, в то же время выступают в качестве крупных географических барьеров. На первом месте стоит распределение теплоты, как фактора, определяющего своеобразие биот широтных зон. Например, преградой на пути распространения деревьев на север в основном является изотерма самого теплого месяца около 10îС. Ниже этой температуры деревья, как правило, расти не могут. Это одна из причин безлесья тундры. На втором – количество осадков и их сезонное распределение. Соотношение тепла и влаги оставляет свой отпечаток на зональных типах биот: виды тропических лесов, живущие в условиях теплого и влажного климата, не расселяются в жаркие и сухие пустыни. Наряду с температурой роль географических барьеров в океане играют изменения солености воды. Отклонение солености от средней океанической (35 î/îî) в сторону опреснения является барьером на пути проникновения многих морских организмов в солоноватые и пресные воды (рис. 3.11).
Существуют организмы, сама природа которых ущербна с точки зрения потенции к завоеванию пространства. Другие, напротив, практически не имеют ограничений к расселению в разные жизненные среды. Так, у иглокожих отсутствует механизм осморегуляции, поэтому их ареал ограничен океаническими водами с полной морской соленостью. Членистоногие же, обретя способность регулировать или защищать внутриклеточное давление, смогли завоевать океан, пресные воды и сушу.
С ландшафтно-экологическими факторами связана внутренняя структура ареала. Ни один вид не образует сплошного покрова. Внутри ареала организмы приуро- чены только к определенным экологическим условиям (местообитаниям). Характер местообитаний контролируется структурой ландшафта. Например, географические ареалы болотных мхов, трав и кустарничков (сфагнума, пушицы, клюквы и др.) и видов таежного мелкотравья и кустарничков (майника, кислицы, черники и др.) в пределах
54

|
|
|
пути расселения |
||
|
|
|
вида встают не |
||
|
|
|
только географи- |
||
|
|
|
ческие и ланд- |
||
|
|
|
шафтно-экологи- |
||
|
|
|
ческие преграды, |
||
|
|
|
но и сопротивле- |
||
|
|
|
íèå, |
которое |
|
|
|
|
встречает ново- |
||
|
|
|
сел со стороны |
||
|
|
|
уже существую- |
||
|
|
|
щих видов. Что- |
||
|
|
|
бы завоевать про- |
||
|
|
|
странство, новый |
||
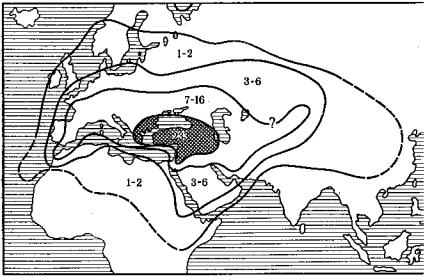
Рис. 3.11. Качественное обилие средиземноморской фауны и флоры и их обеднение с |
вид, должен побе- |
||||
проникновением на восток, по Л. А. Зенкевичу: |
|
дить их в борьбе |
|||
1 – средиземноморская биота; 2 – каспийская биота; 3 – пресноводная биота; |
за существова- |
||||
4 – арктические иммигранты. Цифрами показано количество средиземноморских ви- |
ние. Расширению |
||||
äîâ |
|
|
ареала противо- |
||
|
|
|
|||
тайги совпадают. Однако первые приуроче- |
стоят не только отдельные организмы, но и |
||||
ны к верховым болотам, а вторые к более |
целые сообщества. Например, на границе |
||||
сухим местообитаниям, занятым темно- |
леса и степи внедрению деревьев в степное |
||||
хвойными зеленомошными лесами. Таким |
сообщество препятствует конкуренция с |
||||
образом, экологическая структура ареалов |
мощной дерниной степных трав. |
|
|||
названных видов образует |
|
|
|
|
|
сложное кружево, рисунок |
|
|
|
|
|
которого определяется рас- |
|
|
|
|
|
пределением морфологичес- |
|
|
|
|
|
ких единиц ландшафта – |
|
|
|
|
|
верховых болот и зелено- |
|
|
|
|
|
мошных лесов. Более полно |
|
|
|
|
|
пеструю картину распреде- |
|
|
|
|
|
ления таежных видов внут- |
|
|
|
|
|
ри ландшафта передает про- |
|
|
|
|
|
филь (рис. 3.12). Четко вид- |
|
|
|
|
|
но, что ландшафтно-эколо- |
|
|
|
|
|
гические комплексы контро- |
|
|
|
|
|
лируют характер местооби- |
|
|
|
|
|
таний так, что определенные |
Рис. 3.12. Профиль, иллюстририрующий комбинации растительности и |
||||
виды образуют более плот- |
почв на озерно-ледниковом моренном рельефе в Карелии, по Б. В. Виног- |
||||
ные популяции там, где ус- |
радову: |
|
|
|
|
1– березняки злаково-разнотравные на подзолах маломощных на валун- |
|||||
ловия жизни для них благо- |
|||||
ных супесях; 2 – ельники чернично-долгомошные на подзолах среднемощ- |
|||||
|
|||||
приятны. Это находит отра- |
ных на озерно-ледниковых суглинках; 3 – ельники-брусничные на подзо- |
||||
жение, прежде всего, в рас- |
лах на моренных супесях; 4 – ельники-долгомошники на торфянисто-под- |
||||
пределении показанных на |
золистых почвах на озерно-ледниковых суглинках; 5 – ельники-чернич- |
||||
профиле растительных сооб- |
ные на подзолах среднемощных на моренных супесях; 6 – сосняки-брус- |
||||
ничники на подзолах песчаных на флювиогляциальной морене; 7 – сосня- |
|||||
ществ. |
|||||
ки осоково-сфагновые на торфянистых почвах; 8 – ельники осоково-сфаг- |
|||||
|
|||||
В роли биологических |
новые на торфянистых почвах болот переходного типа; 9 – сосняки сфаг- |
||||
барьеров выступают конку- |
новые на торфяных болотах верхового типа;10 – березняки черничные на |
||||
рентные отношения. На |
подзолах среднемощных супесчаных на моренных супесях |
|
|||
55
3.4. Типы ареалов
Первоначальную область распространения вида называют центром его происхождения. Наряду с этим выделяют ту часть ареала, в которой наблюдается максимальная изменчивость данного вида – наибольшее количество географических форм в пределах вида, видов и подвидов в пределах рода и т. п. Эту часть ареала называют центром многообразия форм (рис. 3.13).
Для возникновения центра многообразия форм требуется время и подходящие условия. Как правило, такой центр тяготеет к более или менее древней части ареала. Если он совпадает с центром происхождения вида, значит, на протяжении его истории условия существования были благоприятными. Но вместе с тем современный центр многообразия форм того или иного вида может не совпадать с центром его происхождения. Так, центр происхождения предковой формы дарвиновских вьюрков лежит на материке, однако наибольшее разнообразие видов наблюдается на Галапагосских островах.
Границы ареала могут быть статичными, расширяющимися или сужающимися. В первом случае устойчивость границ объясняется тем, что вид достиг естествен-
ных границ своего ареала. Границы ареала расширяются, когда молодой вид обладает потенцией к преодолению географических, ландшафтно-экологических и биологических преград. Границы ареала сужаются, когда в силу изменения выше названных условий вид теряет завоеванную территорию (акваторию).
При типизации ареалов следует учитывать их размеры, непрерывность (континуальность) и прерывистость (дизъюнкции).
У наземных организмов выделяются ареалы четырех уровней размерности: континентальные, провинциальные, региональные и локальные. Ареал континентального типа охватывает большие площади одного или нескольких континентов; провинциального занимает большую часть физико-географической или биогеографической провинции; ареал регионального типа ограничен площадью физико-геогра- фического или биогеографического района; локальный тип совпадает с внутриландшафтными морфо-
логическими комплексами – урочищами и фациями (рис. 3.14).
Континуальное (сплошное) распространение обозначено на рисунке нечетными номерами (1, 3, 5, 7), дизъюнкции ареалов – четными (2, 4, 6). Другие типы распространения обозначены буквами А, Б, В. Тип А характеризуется сплошным распространением по всей площади; тип Б состоит из одного большого и одного или нескольких небольших участков, связанных между собой; тип В объединяет множество мелких участков.
Рассмотрим иллюстрации. Виды, распространенные на всех континентах, называют космополитами (что означает “распространенный по всему миру”). Ареал, близкий к типу 1А, имеет папоротник орляк (рис. 3.15). Космополитами являются мно-
56
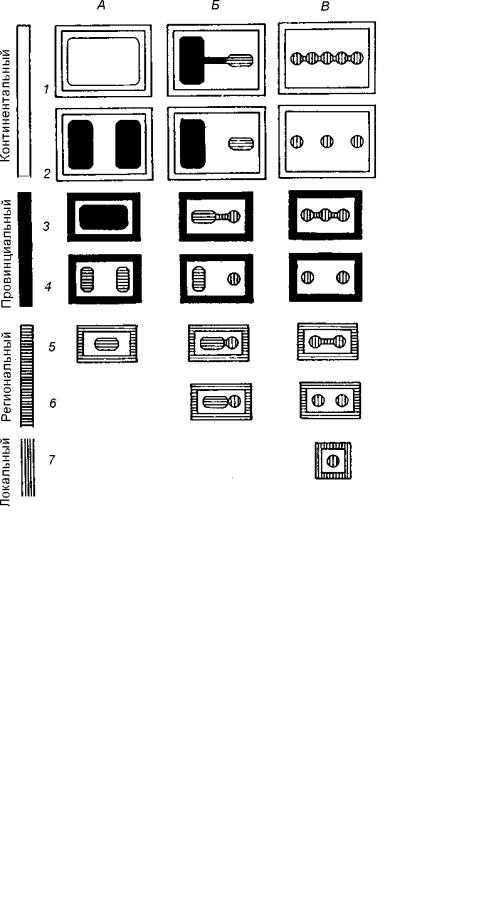
Рис. 3. 14. Основные типы ареалов на суше, по P. Dansereau. Объяснения в тексте
Рис. 3.15 Ареал папоротника орляка (Pteridium aquilinum), по А. И. Толмачеву
57
гие сорняки, связанные в своем распространении с деятельностью человека: мокрица (Stellaria media), крапива (Urtica dioica), одуванчик (Taraxacum officinale). Однако, строго говоря, контрастность экологических условий на Земле не позволяет ни одному виду растений или животных, кроме человека, иметь истинно космополитическое распространение.
Возникновение разорванных ареалов объясняется событиями прошлых геологи- ческих эпох: исчезновением мостов-пере- ходов на суше, дрейфом континентов, горообразованием, изменениями климата и т. п. Ареал вида может быть расчлененным,
èзанимать обширные области на разных континентах, тип 2А (рис. 3.16).
Ареал таежного кустарничка Linnea borealis в основном сплошной, занимающий значительную часть Евразии. Наряду с этим имеются его небольшие обособленные островки в различных местах Средней
èВосточной Европы, на Кавказе, на юге Западной Сибири, на Дальнем Востоке (рис. 3.17). Следовательно, такой ареал можно отнести к типу 2Б.
Рассмотрим историю формирования разорванных ареалов на следующих примерах. Дизъюнкция ареала рода Nepentes объясняется расколом в мезозое материка Гондвана. В результате некогда сплошной ареал оказался разорванным. Ныне виды Nepentes характерны для далеко отстоящих флор Мадагаскара и Юго-Восточной Азии (рис.3.18).
Âверхних поясах гор Центральной и Южной Европы имеются виды, которые отсутствуют на равнинах, но встречаются в Арктике. В данном случае разорванность ареала объясняется похолоданием климата в плейстоцене. Во время ледниковой эпохи в центре Европы царил холодный климат,
èона давала приют растениям и животным, характерным для ландшафтов тундры. В ходе потепления в голоцене эти виды остались в арктических ландшафтах, а южнее были вытеснены в высотные пояса гор, где они смогли выжить и стали ледниковыми реликтами. Так заяц-беляк, основной ареал которого охватывает тундровые и таеж-
ные ландшафты, живет обособленно в Альпийских горах (рис. 3.19). По предложенной выше типологии данный тип ареала можно обозначить индексом 4Б. Этими же причинами вызван европейско-дальневос- точный разрыв ареалов широколиственных лесов. Иллюстрацией может служить ареал спутника широколиственных лесов Hepatica nobilis (рис. 3.20); тип ареала 4А.
Чтобы подчеркнуть определенные особенности ареалов континентальной размерности используют дополнительные термины. Ареалы видов, приуроченных к аркти- ческому поясу северного полушария, называются циркумполярными. Они образуют кольцо вокруг полюса (рис. 3.21). Ареал видов, охватывающих тропический пояс, называется пантропическим (рис. 3.22).
Особый тип образуют биполярные ареалы. Это разорванные ареалы, часть которых расположена в высоких широтах Северного, а часть в высоких широтах Южного полушария (рис. 3.23, 3.24). Возникновению биполярных ареалов способствовало похолодание климата в ледниковую эпоху, когда холоднолюбивые виды Севера смогли преодолеть современный теплый пояс и проникнуть в высокие широты Южного полушария.
Провинциальные и региональные типы ареалов формируются внутри континентов и биоклиматических зон. Например, для европейских древесных пород давно уже установлено, что их распространение на север и восток в наибольшей степени регулируется зимними температурами. Зависимость эта еще ярче отражается в очертаниях ареала европейского падуба (Ilex aquifolium), входящего в состав кустарникого яруса широколиственных лесов (рис. 3. 25); тип ареала 5А.
Примером разорванного ареала региональной размерности (тип 6В) может служить распространение норвежской полыни (рис. 3.26).
Ареалы локальной размерности (тип 7В) характерны для молодых видов, освоивших специфичные местообитания, например дарвиновских вьюрков. Или для реликтовых видов, сильно сузивших свой
58

Т.И уолмачев
.À
suecicaCornus ïî),
едсквш (ого
реА кизилаал
.3 .16
.Ðèñ
Т.И уолмачев
.À
делахпре поЕвразии,
âealis
borLinnaea
ногтаеж ко аустарничк
àëðå
.3 À.17
.Ðèñ
59
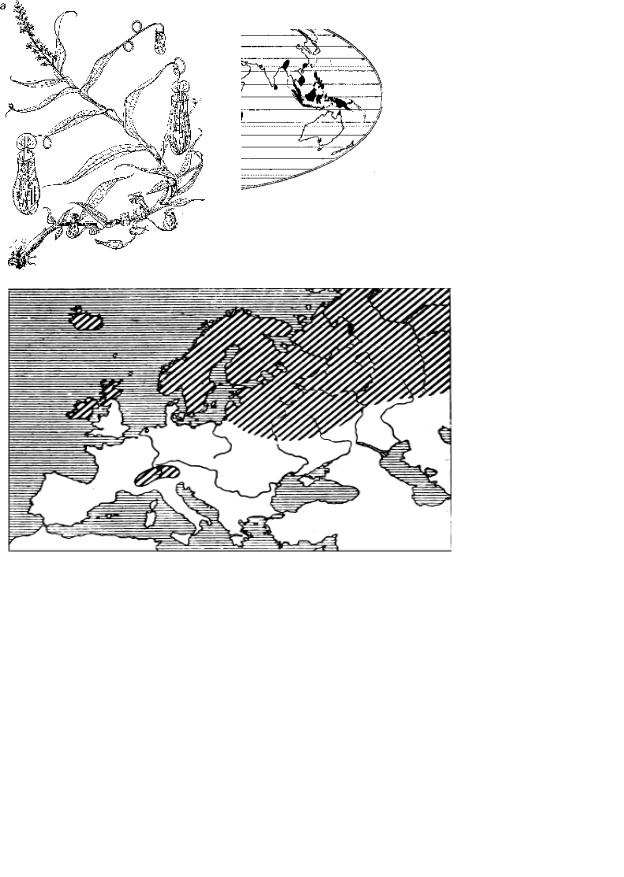
Рис. 3.18. Разорванный ареал рода Nepentes, по R. Good: а – общий вид растения; б – черным цветом залит ареал
Рис. 3.19. Разорванный арк- то-альпийский ареал зайцабеляка в Европе, по Н. А. Бобринскому
Рис. 3.20. Разорванный европейско-дальневосточный ареал Hepatica nobilis, по А. И. Толмачеву
60