

Петров_К_М_Биогеография_СПб_2001
.pdfóÁ îêäû
.Ì .È
Г.А орьевриг иу
.À
óøñ ïîè,
льностиона
зской
П дичеерио закский гон ичееограф
41
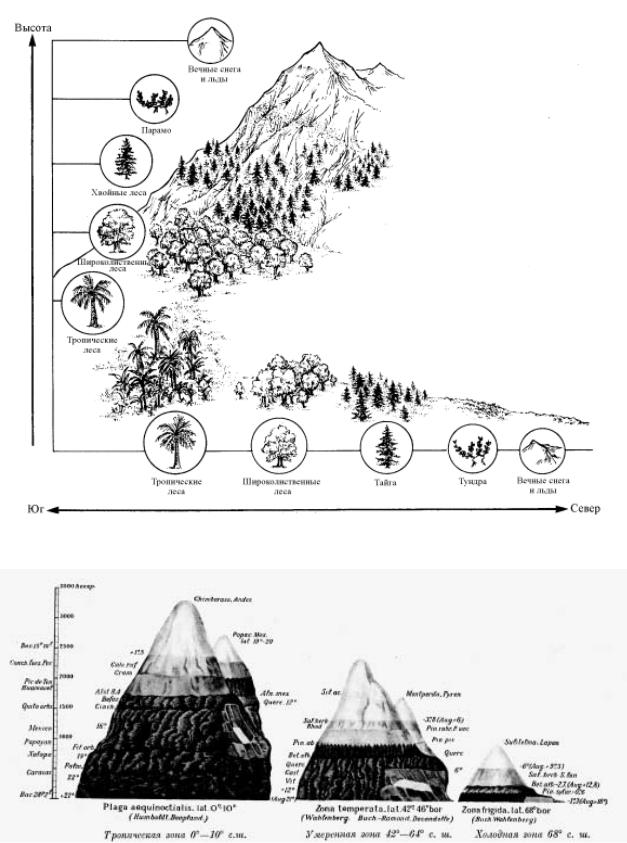
Рис. 2.15. Зеркальное отображение широтных зон в высотных поясах гор, по D. Chiras
Рис. 2.16. Спектры высотной поясности на разных широтах, по А. Гумбольдту:
Наиболее полно высотные пояса представлены на экваторе, на широте 0 – 10î; в умеренных широтах 42 – 46î с. ш. число зон сокращается; в холодной зоне на широте 68î с. ш. их становится еще меньше
42
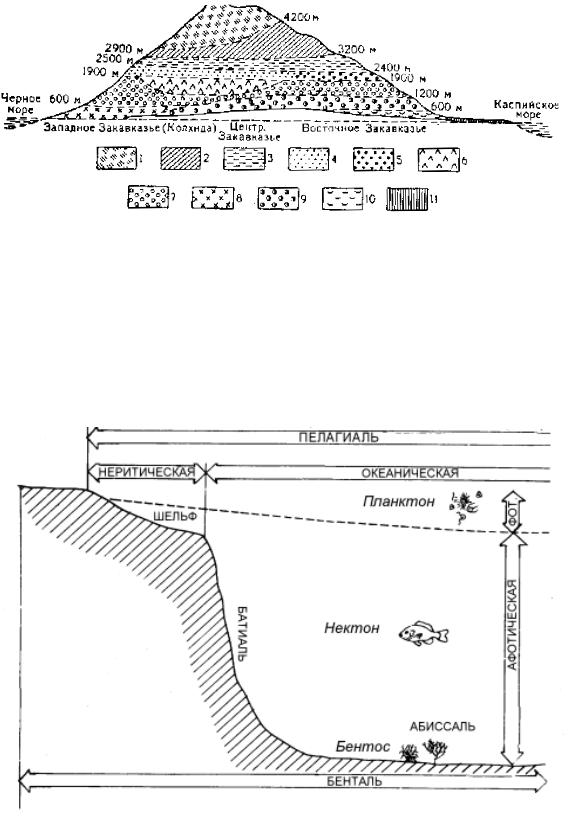
Рис. 2.17. Схема высотных поясов растительности Закавказья, по С. В. Калеснику:
Пояса: 1 – вечного снега и льда: 2 – высокогорной пустыни; 3 – альпийский; 4 – субальпийский; 5 – высокогорных лесов с преобладанием восточного дуба; 6 – елово-пихтовых лесов; 7 – буковых лесов; 8 – смешанных лиственных колхидских лесов; 9 – лесов с преобладанием грузинского дуба; 10 – полупустынь, степей и аридных редколесий; 11 – пустынь
Рис. 2.18. Основные подразделения арены жизни в океане
43
чередование зональных типов ландшафтов определяется изменениями радиационного индекса сухости.
Секторность – универсальная закономерность географической оболочки. Она обусловлена взаимодействием океанов и материков. От соотношения океанических и континентальных воздушных масс зависит степень континентальности климата, количество осадков. Это позволяет разделить материки на физико-географические секторы – крупные региональные единицы, обычно простирающиеся в направлении, близком к меридиональному. В каждом секторе зональность приобретает свою специфику. В приокеанических секторах зональные контрасты сглажены. Для них характерен спектр широтных зон с доминированием лесных типов растительности от тайги до экваториальных лесов. Континентальный спектр зон отличается преобладанием степей, полупустынь, пустынь.
2.3. Подразделения арены жизни, обусловленные действием закона высотной поясности
Высотная поясность, будучи функцией рельефа, обязана своим существованием понижению температуры и увеличению атмосферного увлажнения с высотой. В самых общих чертах вертикальные климати- ческие изменения аналогичны зональным (рис. 2.15). В действительности полного тождества между широтными зонами и высотными поясами не наблюдается. Так, высокогорный пояс парамо в Андах не тождествен зоне тундр; точно так же, как высокогорные луга и леса не тождественны лугам и лесам соответствующих широтных зон.
Высотная поясность начинается с той широтной зоны, в которой находится горная страна. Там, где высота гор позволяет, поясность замыкается вечными снегами и льдами. При этом в высоких широтах снеговая граница опускается до уровня моря;
в жарких странах она поднимается. Выше всего граница снега расположена в сухих субтропиках и тропиках (в Тибете 7 км); в условиях влажного климата на экваторе она опускается до 4,4 км.
Принято говорить о спектрах высотной поясности, которые по-разному проявляются в разных физико-географических условиях. У каждой широтной зоны свой набор высотных поясов. Первым на особенности высотной поясности в зависимости от географической широты указал А. Гумбольдт (рис. 2.16.).
Подобно широтной зональности, высотная поясность бывает двух основных типов: океанического и континентального. В первом случае, в умеренных и теплых зонах лесной пояс начинается на приморской равнине; во втором – на равнине располагаются субаридные и аридные ландшафты (степи, полупустыни, пустыни), а древесно-кустарниковые формации начинаются на известной высоте. На больших горных массивах, на склонах гор разной экспозиции наблюдается асимметрия высотной поясности: спектры высотной поясности выражены по-разному (рис. 2.17).
2.4. Основные подразделения арены жизни в океане
Толща вод Мирового океана представляет самую мощную и обширную часть биосферы – пелагиаль. Ложе океана – бенталь, арена жизни организмов, ведущих донный образ жизни. Пелагиаль подразделяется на верхний, освещенный солнцем фотический слой, и глубоководное царство вечной ночи
– афотическую толщу. В прибрежной, неритической области условия существования отличаются от жизни в пелагиали открытого океана. На морском дне выделяются глубинные пояса, связанные с литоралью,
батиалью и абиссалью (рис. 2.18).
Более подробно подразделения арены жизни в океане рассмотрены в учебном пособии К. М. Петрова (1999).
44

Глава 3
АРЕАЛОГИЯ
Ñовокупность видов растений, встречаю-
щихся на той или иной территории (акватории), образует флору данной территории, совокупность видов животных –фауну. Исторически сложившаяся совокупность живых организмов (флора и фауна) образует биоту. В задачи биогеографии входит районирование биоты, выявление неповторимых флористических и фаунистических комплексов путем анализа особенностей географического распространения систематических таксонов (видов, родов, семейств и т. д.). Поэтому основополагающим разделом биогеографии является учение об
ареалах (ареалогия).
Ареал – это часть земной поверхности или акватории, в пределах которой встречается данный вид организма. Размеры, форма, внутренняя структура, границы ареала определяются историческими, географическими и экологическими факторами.
Методика выявления ареалов достаточ- но проста: на карту наносятся места нахождения видов. Чем полнее проведены наблюдения и точнее определена систематическая принадлежность видов, тем точнее будет картина их географического распространения. Ареалы видов очерчивают наиболее дробные единицы районирования. Чем выше таксон, тем обширнее область его распространения. Ареалы высших таксономических единиц, как правило, охватывают всю сушу, весь океан или всю биосферу.
Работа по картографированию ареалов долгая и кропотливая. Требуются многолетние сборы фактического материала, чтобы, соединив периферийные точки линией, очертить границы ареала. Новые находки, уточнение систематической принадлежности организмов, подчас вносят существенные коррективы в форму ареала, заставляют по новому интерпретировать историю его формирования.
3.1. Формирование ареала
Формирование ареала связано с образованием вида и завоеванием им пространства. Новые виды образуются путем измен- чивости предковых форм, естественного отбора и сохранения наиболее приспособленных, победивших в борьбе за существование. Процесс увеличения биоразнообразия опирается на теорию монофилитической эволюции Ч. Дарвина.
Ч. Дарвин (1809 – 1882)
Согласно этой теории все многообразие видов в крупной таксономической группе растений или животных происходит от одного корня в результате дивергенции – расхождения признаков. На первых этапах дивергенция осуществляется в рамках микроэволюционного процесса, т. е. изменчи- вости существующих организационных признаков. Она носит ярко выраженный приспособительный (адаптационный) характер.
В качестве иллюстрации рассмотрим изменчивость вьюрков с Галапагосских островов, на которую обратил внимание Дарвин (рис. 3.1). На этих островах, располо-
45
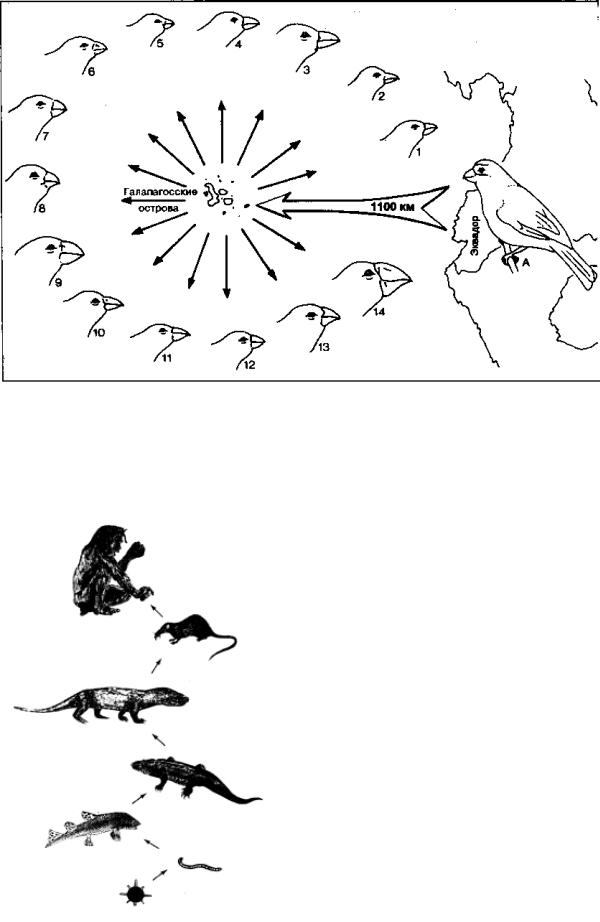
Рис. 3.1. Микроэволюция на примере дарвиновских вьюрков, по Р. Юнкер и З. Шерер:
1 – вьюрок с Кокосового острова; 2 – галапагосский певчий вьюрок; 3 – дятловидный вьюрок; 4 – мангоровый древесный вьюрок; 5 – малый вьюрок; 6 – средний вьюрок; 7 – большой вьюрок; 8 – вьюрок, обитающий на лиственных деревьях: 9 – большой кактусовый вьюрок; 10 – малый кактусовый вьюрок; 11 – остроклювый обыкновенный вьюрок (основной тип); 12 – малый земляной; 13 – средний земляной; 14 – вьюрок толстоклювый, обыкновенный. За исходный тип (A) принят зерноядный земляной вьюрок с Южноамериканского континента
Рис. 3.2. Макроэволюция от простейших до человека, по Р. Юнкер и З. Шерер
женных в Тихом океане южнее экватора в 1100 км к западу от Южной Америки, обитают лишь немногие виды организмов. Среди них заметную роль играют 14 видов вьюрков. Их предком по всей видимости являлась популяция вида, занесенного на один из островов сильным ветром с материка. В результате адаптивной радиации в ходе приспособления к различным условиям существования возникли новые виды, занявшие различные экологические ниши.
Макроэволюция связана с арогенезом (ароморфозом) – крупными анатомо-мор- фологическими и физиологическими изменениями, в результате которых биологический таксон поднимается на принципиально новую, более прогрессивную ступень развития (рис. 3.2). К ароморфным изменениям относятся: появление теплокровности, четырехкамерного сердца, возникновение перьевого покрова и приспособлений к полету и т. п. Примерами дивергентной макроэволюции могут быть млекопитающие и птицы, произошедшие от общего
46

корня – древних пресмыкающихся.
Виды, попавшие в условия географи- ческой изоляции, дают начало ветвям филогенетического древа, все дальше отходящими от основного ствола сначала на уровне микро-, а затем и макроэволюционной дивергенции. Таким образом, формируются неповторимые черты биот, отличающиеся не только по составу видов, но и родов, и семейств.
Каждый вид существует определенное время, его популяция растет, увеличивается площадь распространения. Затем в силу разнообразных причин происходит вымирание особей отдельных видов, их ареалы сокращаются и, наконец, они исчезают. В одних случаях исчезновение вида не прерывает хода эволюции, старый вид вытесняется новым; в других – гибель вида обозначает слепую ветвь филогенетического древа.
Процесс формирования нового ареала, как правило, монотопичен – возникновение вида происходит в одном месте, откуда он начинает завоевывать пространство. Новый вид происходит от одного предка или в результате гибридизации. Эти положения являются ключевыми для понимания истории формирования ареала.
В.И. Вернадский отметил огромную внутреннюю потенцию живого вещества к растеканию по земной поверхности. Он назвал это явление давлением жизни. Оно выражается во “всюдности” жизни,
в захвате ею всякого свободного пространства биосферы. Огромная энергия давления жизни определяется быстротой размножения. Одним из первых количественную модель прогрессии размножения разработал Томас Мальтус.
Вернадский приводит формулу геометрической прогрессии размножения:
2n D = Nn,
где n – число дней с начала размножения;D - показатель прогрессии, который для одноклеточных организмов, размножающихся делением, соответствует числу поколений в сутки; Nn – число неделимых, существующих благодаря размножению че- рез n дней. Характерным для каждого вида является D.
В этой формуле никаких пределов, никаких ограничений ни для n, ни для D, ни для N не содержится. Процесс мыслится бесконечным, как бесконечна сама прогрессия. Растекание живого вещества задерживается только внешними силами. Оно замирает при низкой температуре, при недостатке пищи, при отсутствии места для обитания, из-за конкуренции с другими организмами. Если нет внешних препятствий, всякий вид в характерное для него время может благодаря размножению покрыть весь земной шар, произвести потомство, равное по массе океану или земной коре (табл. 3.1).
Таблица 3.1.Скорость заселения поверхности Зем-
ли, по О. Е. Агаханянцу
Организмы |
|
Скорость заселения |
|
|
|
Бактерии |
|
До 1,8 суток |
Насекомые |
|
203 – 366 суток |
Цветковые растения (клевер) |
|
Около 11 лет |
Ðûáû (êàðï) |
|
Около 12 лет |
Птицы (куры) |
|
Около 18 лет |
Млекопитающие: |
|
|
Крысы |
|
Около 8 лет |
Ñëîí |
|
Около 1000 лет |
|
Вернадский приводит теоретически возможные скорости растекания жизни. Она колеблется от 33 100 см/с у бактерий, т. е. близка к скорости звука, до долей сантиметра в секунду у крупных млекопитающих – у индийского слона, например, 0,09 см/с.
Расселению организмов способствуют различные приспособления их зачатков (диаспор): при половом размножении растений – спор, плодов, семян, при вегетативном – луковиц, клубней, корневищ, ползу-
47
чих побегов и т. д. У животных роль диаспор могут играть яйца, личинки, цисты.
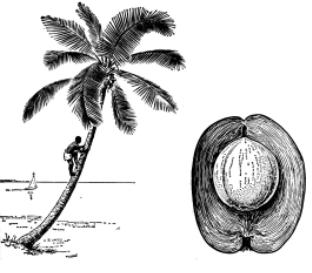
Плоды и семена большинства водных, а равно и значительного количества наземных растений легко переносятся текучими водами, чем облегчается расселение растений вниз по течению рек. Многие плоды и семена могут переноситься морскими те- чениями. Например, ареал кокосовой пальмы (Cocos nucifera) охватывает берега тропических островов, разбросанных по акваториям Тихого и Индийского океанов. Кокосовые орехи, попадая в воду, разносятся морскими течениями. Они хорошо приспособлены к длительным морским путешествиям. Своей плавучестью они обязаны заполненной воздухом и непроницаемой для соленой морской воды волокнистой наружной оболочке (рис. 3.3). Орехи не теряют способности к прорастанию после плавания в море в течение 3 – 4 месяцев. За это время они могут быть перенесены на расстояние до 5000 км.
Легкость спор и мелких семян множества видов обеспечивает возможность переноса их на более или менее значительное расстояние даже слабыми токами воздуха. У растений с более тяжелыми семенами (или плодами) возможность переноса по воздуху часто возрастает благодаря “летательным приспособлениям” – волосистым придаткам, разного рода хохолкам, перепон- чатым лопастям, “крылышкам” и т. д., под- час обладающим довольно сложным устройством (рис. 3.4).
Перенос зачатков растений при помощи ветра может осуществляться не только по воздуху: опавшие на землю плоды и семена, при наличии у них летучек, могут перегоняться ветром по поверхности поч- вы; семена деревьев, опавшие зимой на затвердевшую поверхность снежного покрова, перегоняются, скользя по ней, особенно легко. Упав на поверхность воды, снабженные летучками семена, долго не тонут и под напором ветра легко передвигаются по поверхности водоемов.
У многих растений семена или плоды переносятся животными благодаря разного рода прицепкам, которыми они прикреп-
Рис. 3.3. Кокосовая пальма, по: “Жизнь растений”: Общий вид пальмы на берегу океана и разрез плода, на котором видна толстая волокнистая оболочка, обеспечивающая хорошую плавучесть ореха
ляются к шерсти или другим покровам (рис. 3.5). То же может достигаться и при помощи клейких выделений.
Семена растений, плоды которых поедаются животными, переносятся последними в кишечнике и попадают на поверхность почвы с пометом. Перенос семян на значительные расстояния птицами играет особо важную роль для расселения растений. Заметим, что вместе с экскрементами в почву вносятся питательные вещества. А. Г. Воронов отмечает, что на сфагновых болотах, почти лишенных питательных веществ, семена голубики прорастают только в том случае, если они попали сюда с экскрементами птиц. Семена многих растений обретают всхожесть только после того как пройдут через желудочный тракт животного. Бывает и так, что определенные животные делают для себя запасы питательных семян, но используют их не полностью или просто теряют часть своих “складов”, которые тогда и превращаются в рассадники растений соответствующего вида.
Большую роль в распространении растений играет человек. Например, растения вводились в культуру первоначально в центрах своего происхождения, а затем начи- нали культивироваться широко по земному шару. Нередко в новых районах растения чувствуют себя даже лучше и завоевывают большие территории, чем на родине. В ка-
48

Рис. 3.4. Плоды и семена, распространяемые ветром, по К. Шефферу:
1 – одуванчик и его плоды; 2 – плоды чертополоха; 3 – семена сосны; 4 – плоды клена; 5 – соплодие липы; 6 – плоды вяза; 7 – плод граба; 8 – плоды березы
Рис. 3.5. Плоды, распространяемые животными, по К. Шефферу:
1 – соплодие лопуха; 2 – побег подмаренника цепкого с плодами; 3 – одиночный плод череды
49

честве примеров можно привести плантации кофе (родина Эфиопия), бананов и цитрусовых (родина Юго-Восточная Азия) в Южной Америке; картофеля, кукурузы, подсолнеч- ника (родина Центральная и Южная Америка) в Европе и Азии. Многие вредные растения и сорняки разносятся человеком изза неосторожности.
Ярким примером формирования нового ареала является история быстрого расселения в Европе американского по происхождению водного растения элодея (Elodea canadensis). Она была завезена в Европу в 1836 г. Далее ее распространение шло уже помимо воли че- ловека (рис. 3.6). Элодея так быстро заполняла водоемы, что это дало повод назвать ее “водяной чу- мой”.
В способах распространения неподвижных или
мало подвижных животных много общего с растениями. Многие виды пауков летят по воздуху благодаря выпускаемой ими паутинной нити. Ураганы и смерчи переносят насекомых и птиц. Сами животные разносят своих паразитов. Водоплавающие птицы на своих лапках переносят диаспоры растений и беспозвоночных, земноводных и рыб, способствуя обогащению фауны водоемов.
Особенно велика роль морских тече- ний в переносе диаспор животных. Многие донные беспозвоночные (моллюски, иглокожие, кишечнополостные, черви, членистоногие) ведут сидячий образ жизни, прирастая к субстрату, или передвигаются по дну в пределах ограниченной площади. Однако их личинки проходят планктонную стадию развития в поверхностной толще
воды. Здесь они подхватываются течениями и переносятся на многие тысячи миль.
У планктонных организмов развились замечательные приспособления для того, чтобы без лишней затраты усилий держаться в толще воды. При помощи различных выростов, щетинок они приобретают большую поверхность по сравнению с массой тела (рис. 3.7). Кроме того, планктонные организмы содержат капельки жира, которые одновременно служат запасным веществом и облегчают массу тела в воде.
Подвижные формы животных расселяются активно. Два главных фактора определяют их стремление к перемещению: инстинкт продолжения рода и поиски пищи. Для многих птиц районы, богатые пищей и пригодные для выведения птенцов в летний сезон, неблагоприятны для жизни в зимний.
50