

Петров_К_М_Биогеография_СПб_2001
.pdfгрунтовых вод, питающих живой раститель- |
формированию на ней кочковато-бугристо- |
||
ный покров, постепенно уменьшается: уже |
го болота (БГЦ-46). Растущий |
выпуклый |
|
не все корни достигают минерального грун- |
торфяник вызывает заболачивание примы- |
||
та. В снабжении растений водой все боль- |
кающего к нему леса. Выпуклый, кустарни- |
||
шую роль начинают играть атмосферные |
ково-лишайниковый, сухой, мерзлый торфя- |
||
осадки. Низинные болота вступают в ста- |
ник (III). Сток идет только в одном направ- |
||
дию переходных мезотрофных (средних по |
лении – с торфяника в сторону леса. Рост |
||
богатству минерального питания) болот. |
торфяника вверх прекращается, он распол- |
||
Это преимущественно сфагновые болота с |
зается вширь, вследствие чего резко увели- |
||
мощностью торфа более 1 м. |
|
чивается скорость заболачивания и гибели |
|
При значительном повышении повер- |
лесов, а также формирования многолетне- |
||
хности болота, приводящем к полному от- |
мерзлых грунтов |
|
|
рыву корневых систем растений от грунто- |
Сфагновые мхи являются растениями- |
||
вых вод, живой растительный покров на- |
эдификаторами, полностью |
контро- |
|
чинает питаться преимущественно атмос- |
лирующими весь комплекс экологических |
||
ферными осадками, бедными минеральны- |
условий: гидрологический и газовый режи- |
||
ми солями, и болото переходит в стадию |
мы, температуру корнеобитаемого горизон- |
||
верхового сфагнового олиготрофного (бед- |
та, образование форм мезо- и микрорелье- |
||
ного питания) болота. Мощность торфа |
фа, процессы почвообразования, точнее |
||
сфагновых болот в среднем 3-4 м. |
|
формирования торфяной залежи. |
|
Рассмотрим процесс формирования |
Сфагнум впитывает атмосферные осад- |
||
верхового болота в подзоне северной тай- |
ки, как губка. Среди клеток, составляющих |
||
ги. Обратим внимание на то, что |
|
|
|
вместе с торфяником возникает |
|
|
|
толща многолетнемерзлых грунтов |
|
|
|
(ðèñ. 9.100). |
|
|
|
Нарастание мохово-торфяни- |
|
|
|
стого слоя ведет к ухудшению |
|
|
|
гидротермического и аэрациопно- |
|
|
|
го режимов почвы, в которой не |
|
|
|
могут существовать корни деревь- |
|
|
|
ев (I). На месте низины поднима- |
|
|
|
ется выпуклый торфяник, с кото- |
|
|
|
рого сметается снег, что способ- |
|
|
|
ствует интенсивному промерза- |
|
|
|
нию торфяника и формированию |
|
|
|
под ним вечной мерзлоты мощно- |
|
|
|
стью в несколько десятков метров |
|
|
|
(II). Между торфяником и лесом об- |
|
|
|
разуется понижение, в которое сте- |
|
|
|
кает вода не только со стороны |
|
|
|
леса (расположенного в верхней |
|
|
|
части равнины), но и с торфяника. |
|
|
|
Это ведет к формированию пере- |
|
|
|
увлажненной полосы на границе |
|
|
|
между торфяником и лесом – мо- |
Рис. 9.100. Схема заболачивания низины и роста торфяника в подзо- |
||
хово-осокового болота (БГЦ-5), ко- |
не северной тайги, по В. В. Крючкову: |
|
|
торое, в свою очередь, подтапли- |
1 – направление стока воды; 2 – граница многолетнемерзлых (веч- |
||
вает и заболачивает полосу, при- |
номерзлых) грунтов; 3 – погребенные в торфяниках стволы деревь- |
||
ев; 4 – торфяные кочки; 5 – торфяники; 6 – бугры пучения с ново- |
|||
мыкающую к лесу, и способствует |
образованиями мерзлоты. Пояснения в тексте |
|
|
211
его листья и стебель, имеются крупные мертвые клетки с отверстиями в оболочке, которые легко наполняются водой. В сухой сезон года головки сфагнума могут высыхать до воздушно-сухого состояния. Однако если вырвать пучок мха из болотной кочки, мы увидим, что в своей нижней части он напитан водой. Сжав сфагнум в руках, можно выдавить из него воду.
Погрузим руку в толщу сфагнума. Рука свободно уходит в мох. Раздвинув куртину, видим, что стебельки мха внизу теряют свою индивидуальность, все сливается в сплошную коричневую массу полуразложившихся частей болотных растений – это торф. Почвы в обычном понимании здесь нет. Все растения укореняются или в еще живом сфагнуме или в торфе.
Ощущается быстрое понижение температуры с глубиной. Если температура воздуха 25 °С, то на глубине 20 см она может уменьшиться вдвое, а на глубине 50 см опуститься до 5 - 6 °С. Резкое снижение температуры с глубиной объясняется плохой теплопроводностью как живого сфагнума, так и торфа.
Торфяные сфагновые болота характеризуются высокой степенью кислотности, зна- чительно более высокой, нежели почвы хвойного леса. Низкая температура корнеобитаемого горизонта вместе с высокой кислотностью, плохой аэрацией и малым содержанием питательных минеральных солей создают весьма специфические условия существования болотных растений.
Прежде всего они страдают от физиологической сухости. Несмотря на то, что корнеобитаемый горизонт насыщен влагой, она с трудом всасывается корнями. Недостаток минерального, прежде всего азотного питания, физиологическая сухость являются причиной ксероморфизма болотных кустарничков: мелколиственности, жестколистности, образования плотной кутикулы, чешуек, волосков, глубокого расположения устьиц.
Растения холодных и влажных местообитаний образуют особую экологическую группу – психрофиты. Они характерны для растительного покрова тундровых биомов.
Таким образом, облик верховых болот таежной зоны как бы переносит нас в расположенный в арктических широтах тундровый ландшафт. Как для верхового болота, так
èдля тундры, кроме ковра из мхов и лишайников, характерны мелкие кустарнички: подбел (Andromeda polifolia), вереск обыкновенный (Calluna vulgaris), багульник болотный (Ledum palustre), два вида клюквы (Oxycoccus), голубика (Vaccinium uliginosum), болотный мирт (Chamaedaphne calyculata) и др. Это все вечнозеленые рас-
тения, относящиеся к одному семейству вересковых (Ericaceae).
Любопытны флорогенетические связи болотных кустарничков. Их родина – горы тропических широт Юго-Восточной Азии. На это обратил внимание русский географ А. Н. Краснов, исследовавший в начале века растительность высотных поясов островов Зондского архипелага. Вот как описывает он свои впечатления о растительности субальпийского пояса одного из вулканов на о.Ява: “... любопытно, что дошедшие до вершин формы, слагаются в ассоциации, большей частью составленные теми же родами или семействами, которые идут далеко на север
èобразуют зону тундры. Род Agаpetes заменяет нашу голубику. Это Ericaceae – по облику его можно назвать тропическим деревом голубикой... Его плоды и цветы так напоминают чернику, что вы сразу признаете в нем Vaccinium, а черные губы ваших спутников-малайцев напомнят вам ваши детские прогулки по болотам и лесам родины” (1987, с.315).
À.Н. Краснов делает вывод, что “флора сфагновой тундры, защищенная от мороза снежным покровом, ... есть прямой продукт вырождения тропической растительности в полярную под влиянием понижения температуры при сохранении прочих условий, то есть влаги и, возможно, меньшего колебания температуры”.
Тундровый облик растительности верховых болот хранит следы послеледниковья. Сфагновые болота начали формироваться по периферии отступавшего ледника около10 тыс. лет назад. Специфические условия существования растительности на вер-
212

ховых болотах позволили им сохранить свой тундровый облик в таежной зоне до наших дней.
Из древесных пород на верховых болотах растет преимущественно сосна (Pinus sylvestris), образующаяздесьрядэкологических форм, резко отличающихся от высокоствольных деревьев, растущих в лесу на минеральном грунте. Болотная сосна никогда не достигает больших размеров: мутовки ветвей близко расположены одна к другой, что указывает на малый годичный прирост. Возраст сосенки высотой около метра может достигать 50
– 70 лет. На торфяниках с быстрым приростом сфагнума (3 - 4 см в
год) встречается своеобразная кустарниковая форма сосны, у которой на поверхности торфяника видны только молодые побеги, покрытые хвоей.
На поверхности мохового покрова растет росянка (Drosera rotundifolia) – небольшое травянистое растение с розеткой прикорневых листьев. Это удивительное растение заинтересовало Ч. Дарвина. Поставив ряд экспериментов, он установил, что ввиду недостаточного минерального питания росянке необходима белковая пища. Поверхность листа росянки покрыта волосками. На каждом волоске имеется капля жидкости, которая на солнце блестит, как роса. Под действием сока, выделяемого волосками, росянка переваривает попавших на лист мелких насекомых. Питаясь таким образом, росянка получает с белками недостающий ей азот. Содержащиеся в росянке элементы минерального питания постепенно переходят в торф, способствуя повышению его плодородия.
Растительности крупных массивов верховых болот присуща ярко выраженная комплексность, обусловленная хорошо развитым болотным микрорельефом (рис.
9.101). В образовании микрорельефа болота (а также в значительной части мезорельефа) большую роль играет сама растительность.
Верховое болото имеет выпуклую форму (вид перевернутой тарелки), поэтому более обводненными являются его центральная и периферийная части. Сильно обводненная окраинная часть болота занята осоковыми комплексами с шейхцерией (Scheuchzeria palustris). Вместе с шейхцерией произрастают осока топяная (Carex limosa), сабельник (Comarum palustre), вахта (Menyanthes trifoliata). Встречаются кустарники: береза карликовая (Betula nana), ива (Salix myrtilloides) и др. Из сфагновых мхов обычны Sphagnum fallax, Sph. riparium, образующие рыхлый моховой покров. Выше, в нижней половине склона с наиболее активным стоком, развит окраинный сосно- во-кустарничково-сфагновый комплекс. Густой кустарничковый ярус образуют карликовая береза, багульник, голубика, вереск и др.
Еще выше, в верхней половине склона, развивается грядово-мочажинный комплекс. Вытянутые поперек склона гряды чередуются с узкими обводненными моча-
213
жинами. Они резко отличаются друг от друга по структуре и видовому составу. На грядах, сложенных сфагновыми мхами, образуется плотный покров Sphagnum fuscum, Sph. magellanicum, растут болотные кустарнички, сильно угнетенные формы сосны (Pinus sylvestris f. wilkommii). В мочажинах обычны сфагновые мхи, отличающиеся рыхлыми дернинами (Sphagnum cuspi-datum, Sph. balticum, Sph. fallax).
При переходе от склона к более выровненной верхушке сфагнового торфяника возрастает обводненность; грядово-мочажин- ный комплекс сменяется грядово-озерко- вым, в котором отдельные мочажины заполнены водой и принимают вид узких и длинных озер.
На плоской вершине верхового болота сильно увеличивается обводненность. Плоские гряды едва возвышаются над широкими, очень топкими мочажинами, заросшими сфагнумами. На повышениях – Sphagnum fuscum, печеночники, местами пятна лишайников, редкие кустарнички, пушица, осоки, а также единичные сосенки высотой 30 – 50 см (Pinus sylvestrisf. pumila), у которых все стволики погружены в торф и возвышаются только концы ветвей. Местами в центре болота расположены озера, к которым часто невозможно подойти из-за большой топкости и обводненности берегов.
В Неарктической области хвойные леса имеют свои отличительные особенности. Они занимают в Северной Америке очень большие территории: большую часть полуострова Лабрадор, значительные части Аляски и широкую полосу, связывающую эти два полуострова.
Вдоль берегов хвойные леса продвигаются далеко на юг: на западе вдоль тихоокеанского побережья, особенно по горным цепям до самой Центральной Америки (Скалистые горы, Каскадные, СьерраНевада и др.); на востоке по приатланти- ческой прибрежной равнине до Мексиканского залива. Здесь они образуют особые формации, входящие в состав субтропических биомов.
По сравнению с европейскими леса Неарктики отличаются большим разнообразием хвойных пород: многочисленны виды сосны, ели, пихты, лиственницы, можжевельника и пр. Кроме того встреча- ется Tsuga, Pseudotsuga, Thuja. Основной причиной этого богатства являются благоприятные условия миграций во время ледникового периода: все эти породы свободно отступали в убежища на юг, а затем снова возвращались после таяния ледника.
Как и в Европе, в более холодных частях Северной Америки хвойные широко распространены на равнинных территориях, в более же южных частях они переходят в горы.
Наиболее далеко на север заходит белая ель (Picea alba), а также лиственница (Larix americana); на севере Канады преобладающее дерево хвойных лесов сосна (Pinus banksiana), к которой присоединяются некоторые ели, бальзамическая пихта (Abies balsamea), лиственница и мелколиственные - осина, береза.
На Аляске растут леса ситхинской ели
(Picea sitchensis), цуги (Tsuga heterophylla, Т. mertensiana), аляскинского “кипариса” (Chamaecyparis nutkatensis).
В области Великих озер и реки св. Лаврентия леса состоят главным образом из веймутовой сосны (Pinus strobus), к которой присоединяются туя (Thuja occidentalis), цуга (Tsuga occidentalis). Вдоль восточного побережья хвойные леса далеко спускаются к югу и, например, в штатах Вирджиния и Каролина леса состоят главным образом из сосны (Pinus rigida, P. taeda).
На западном побережье хвойные леса представлены особыми видами: так, для Британской Колумбии наиболее характерны туя, дугласова пихта, сосна (Pinus murreyana) и др.
Животный мир Неарктики имеет много общего с таежным биомом Палеарктической области. Отличия прослеживаются на видовом, реже на более высоком таксономическом уровне (рис. 9.102.). Своеобразие таежной фауне Неарктики придают древесный дикобраз, ондатра, скунс; панамериканские формы – колибри, пума.
214

Рис. 9.102. Характерные представители таежной фауны Неарктической области Голарктического царства, по А. Г. Воронову:
1 – енот; 2 – гремучая змея; 3 – голубая сойка; 4 – снежный козел; 5 – колибри; 6 – толсторогий баран; 7 – кардинал; 8 – воротничковый рябчик; 9 – овцебык; 10 – бобр; 11 – древесный дикобраз; 12 – бурундук; 13 – северный олень; 14 – вапити; 15 – ондатра; 16 – лось
215
9.2.3. Широколиственные леса Северного полушария
Широколиственные леса располагаются в приокеанических секторах умереннотеплого пояса Европы, Восточной Азии и Северной Америки. Основу биомов образуют летнезеленые (листопадные) широколиственные породы деревьев. Такие леса называют еще неморальными, а саму зону широколиственных лесов неморальной зоной. В Северном полушарии она полностью принадлежит Голарктическому царству.
Неморальная зона отделена от тайги переходной зоной смешанных хвойно-ши- роколиственных лесов. Подобные зональные типы биомов не характерны для умеренного пояса Южного полушария.
Как уже говорилось, по мере перехода в низкие широты, где климат становится более теплым, все отчетливее проявляется роль увлажнения, и закон географической секторности в распределении зональных типов биомов находит здесь свое более яркое, чем в холодном поясе, выражение. Так почти в одном интервале широт, но на разном удалении от океана формируются биомы широколиственных лесов, степей и пустынь (см. рис. 2.14).
Летнезеленые леса одеты листвой лишь в благоприятное летнее время года, осенью листья опадают и зимой деревья стоят без листьев. Почки на ветвях деревьев покрыты почечными чешуями, а стволы толстой коркой – это защита не только от мороза, но и от зимней засухи, когда вода замерзает и становится мало доступной для метаболизма.
Широколиственные леса отличаются от тайги богатством видового состава и сложной структурой. Наиболее богаты леса Северной Америки и Восточной Азии, поскольку здесь прослеживается их тесная связь с арктотретичной флорой плиоцена (см. гл. 3).
Палеарктика. В Евразии биомы широколиственных лесов тяготеют к океани- ческим секторам Атлантического и Тихого океанов. Широколиственные леса занимают большую часть Западной Европы; в Во-
сточной Европе, где заметно усиливается континентальность климата, леса выклиниваются. Здесь их северная граница проходит приблизительно по линии Калининград – Москва – Уфа, а южная – по линии Кишинев – Êèåâ – Òóëà – Уфа. Таким образом, область распространения этих лесов на Европейском континенте напоминает по форме треугольник, вершиной обращенный к Уралу. В западной части ареала широколиственные леса располагаются к северу от биомов средиземноморского типа; в восточ- ной – к северу от степей.
Соотношение тепла и влаги в неморальной зоне близко к оптимальному. И хотя здесь так же, как в тайге, выражены все че- тыре времени года, и зимой происходит спад биологической активности, продолжительность вегетации удлиняется до 6 -7 месяцев против 4 -5 в бореальной зоне.
В Западной Европе биом широколиственных лесов относительно однороден и образует сплошную зону, ширина которой местами достигает 1500 км (от Южной Скандинавии до северной части Пиренейского п-ова). Температура самого теплого месяца 15 – 18 îС на севере и 18 – 22 îС на юге. Зимы мягкие. Средняя температура января от -5 îС до положительной 4 – 7 îС. Длительность безморозного периода 150 – 200 дней. Среднее годовое количество атмосферных осадков порядка 600 – 800 мм. Испаряемость примерно равна этой вели- чине.
Европейские широколиственные леса намного беднее по составу и структуре, чем биомы данного типа, распространенные в Восточной Азии и Северной Америке. Эта бедность есть следствие четвертичных оледенений. Сохранению в этот период богатой третичной лесной растительности на юге в убежищах препятствовали альпийские ледники и Средиземное море. И все же состав и ярусная структура европейских широколиственных лесов намного богаче и сложнее, чем у таежных биомов. В их сложении, как правило, принимают участие три яруса – древесный, в котором различа- ется до трех подъярусов, кустарниковый (до двух подъярусов) и травяной (с двумя - тре-
216
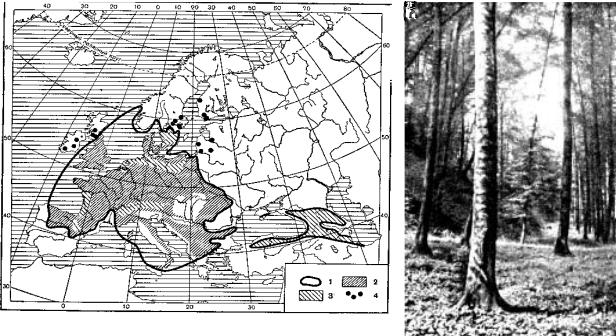
Рис. 9.103. Ареал буковых лесов, по В. В. Алехину:
1 – границы ареала; 2 – области наибольшего распространения;3 – области меньшего распространения; 4
– искусственные посадки; справа – буковый лес
мя подъярусами). Обильная опавшая листва образует мощный слой подстилки. Моховой ярус выражен плохо.
Умеренно-теплый и влажный климат, активная деятельность почвенных организмов, грибов и животных, способствуют быстрому разложению опавших листьев и накоплению гумуса. Господствующий тип почв – бурые лесные.
Лесообразующими породами служат четыре вида дуба (основные – черешчатый
Quercus robur и скальный Q. petraea), бук лесной (Fagus sylvatica), а также ясень обыкновенный (Fraxinus excelsior), граб обыкновенный (Carpinus betulus), несколько видов липы (Tilia), клена (Acer), вяза (Ulmus). В Северной Америке и Восточной Азии по- чти все перечисленные рода деревьев представлены десятками, а дуб и клен даже сотнями видов.
Несмотря на такой сравнительно небольшой набор эдификаторов, европейские широколиственные леса представлены довольно многочисленными вариантами – зональными, провинциальными, высотнопоясными, эдафическими.
В материковой части приатлантического сектора в его северной подзоне распространены буковые леса (рис. 9.103). По составу и структуре это наиболее бедные
леса. Часто в них развит только один древесный ярус. Почва, покрытая толстым слоем опавших листьев, почти лишена яруса трав. В Южной и Центральной Европе буковые леса перемещаются в предгорья и на склоны гор. Здесь же произрастают каштан (Castanea sativa), клен ложноплатановый (Acer pseudoplatanus), липа крупнолистная (Tilia platyphyllos). В приатлантических лесах встречаются вечнозеленые виды – падуб остролистный (Ilex aquifolium), морозник (Helleborus), плющ (Hedera helix).
Для равнин центральноевропейского сектора типичны мезофильные леса из дуба черешчатого (Quercus robur), которому сопутствуют липа сердцелистная (Tilia cordata), клен платановидный (Acer platanoides), граб, ясень. В подлеске присутствуют лещина (Corylus avellana), виды боярышника (Crataegus), бересклет бородавчатый (Euonymus verrucosa), волчье лыко (Daphne mezereum) и другие кустарники. Для травяного покрова характерны подснежники – виды, которые успевают закон- чить вегетацию до облиствения деревьев, например хохлатка полая (Corydalis cava), гусиный лук (Gagea lutea), ветреница дубравная (Anemone nemorosa) или отцвести – сочевичник весенний (Lathyrus vernus), медуница лекарственная (Pulmonaria
217

officinalis), пролесник многолетний (Mercurialis perennis) и др. Из злаков типи- чен перловник (Melica).
Травянистые растения широколиственных лесов, имеющие широкие нежные листовые пластинки, относят к экологической группе дубравного широкотравья. Некоторые из них проникают в таежные биомы, где их называют неморальными элементами. В частности, они составляют характерную черту сложных (неморальных) ельников.
Âюго-восточной части Центральной Европы (внутри горной дуги Карпат) происходит некоторое усиление сухости и континентальности климата при повышенной теплообеспеченности. В широколиственных лесах начинают доминировать дубы австрийский, или цер (Quercus cerris), и венгерский (Q. frainetto), намечая переход, с одной стороны, к субсредиземноморским ландшафтам, а с другой – к лесостепным.
ÂВосточноевропейских широколиственных лесах состав пород меняется. Здесь нет бука, дуба скального, клена явора (Acer pseudoplatanus). Граб распространен только до левобережья Днепра. Ясень не переходит за Волгу. Основной эдификатор дуб черешчатый (Quercus robur) - дерево, способное переносить летнюю засуху и зимние
морозы. Возраст отдельных деревьев может достигать 2000 лет. Их по праву называют лесными патриархами (рис. 9.104). На западе в качестве соэдификатора выступает граб, к востоку усиливается роль липы.
Животный мир широколиственных лесов намного богаче, чем в таежных биомах. Типичные представители показаны на рис. 9.105. Есть виды общие с фауной тайги. Из млекопитающих – это лесная куница, лесная кошка, рысь, черный хорек, барсук, белка, заяц-русак, заяц-беляк, еж. В горных лесах встречается медведь.
Небольшая мощность снежного покрова или даже полное его отсутствие, обилие растительного корма благоприятствуют распространению копытных – зубра, благородного оленя, косули, кабана. Из птиц можно отметить дятла, синицу, дрозда, зяблика и др. Почва широколиственного леса отличается высокой насыщенностью фауной беспозвоночных – насекомыми, дождевыми червями.
Важную часть пищевого рациона зверей и птиц составляют зеленая растительная масса, сочные плоды, семена, желуди и лесные орехи (рис. 9.106).
В средние века европейская зона широколиственных лесов начала активно ос-
Рис. 9.104. Ареал дуба черешчатого, по В. В. Алехину:
Точками обозначена граница ареала ясеня; справа – опушка дубового леса, по П. Е. Васильковскому
218

Рис. 9.105. Характерные представители фауны европейского широколиственного леса. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – лесная куница; 2 – черный хорь; 3 – садовая соня; 4 – полчок; 5 – иволга; 6 – квакша; 7 – орешниковая соня; 8 – зеленый дятел; 9 – дубонос; 10 – европейская неясыть; 11 – белка; 12 – зубр; 13 – европейская косуля
219

лое и дождливое. На материке средняя температура самого холодного месяца - 16 îС; самого теплого около 20 îС. За год выпадает примерно столько же осадков, сколько и в европейских широколиственных лесах (500 – 700 мм), но по сезонам они распределяются крайне неравномерно. Большую часть осадков приносит летний муссон; всю осень и зиму, с октября по апрель, - сухой сезон. В январе – феврале выпадает всего 5 – 10 мм, снежный покров – маломощный. На островах климат значительно мягче, контраст сезонов сглажен. Температура самого холодного месяца около -2 îС. Годовая сумма осадков 1100 – 1800 мм, что более чем в два раза превышает
количество осадков на материке. Зимы снежные.
Леса многоярусные, увитые лианами (рис. 9.107).Широколиственные породы деревьев представлены видами дуба, клена, ясеня, липы, ильма, орехом маньчжурским (Juglans mand-schurica) и др. А также рядом видов, отсутствующих в Европе: софора
(Sophora japonica), гледичия (Gleditschia sinensis), айлант (Ailanthus glandulosa), магнолия (Magnolia kobus), камелия (Camelia japonica), бархат амурский (Phellodendron amurense) и др. Многие из названных растений культивируются в других странах как декоративные.
Очень богат подлесок, состоящий из жимолости (Lonicera), сирени (Syringa), рододендрона (Rhododendron), аралии (Aralia elata) и др. Большинство этих родов представлено многочисленными видами, отсутствующими в Европе. Помимо обильного подлеска, хорошо представлены лианы: амурский виноград (Vitis amurensis), актинидия (Actinidia kolomikta), лимонник
220