

Петров_К_М_Биогеография_СПб_2001
.pdfЭлементарные эталоны – вырезки из аэрофотоснимков, характеризующие изображение четко выделяющихся объектов и явлений. Они систематизируются в виде тематических таблиц и могут располагаться в порядке, отражающем классификационные подразделения структурных единиц ландшафта. Элементарные эталоны оформляются в виде легенды и прикладываются к материалам съемки.
Эталонный профиль представляет собой полоску изображения, на которой отобразилось характерное сочетание сопряженных природных комплексов. Ему сопутствует детальный ландшафтный профиль.
Составные эталоны комбинируются из фрагментов снимков, расположенных по линии регионального профиля, пересекающего ландшафт в направлении характерного изменения ландшафтно-экологических условий. Фрагменты снимков приближаются по форме к элементарным эталонам. Они отображают наиболее существенные изменения изображения в выбранном направлении.
Значение эталонов состоит в том, что с их помощью осуществляется камеральное дешифрирование дистанционных изображений новых, непосещенных территорий методом экстраполяции дешифровочных признаков. Достоверность экстраполяции определяется контрольными проверками.
Внутриконтурная экстраполяция применяется в пределах одного контура изображения. Размер ключевого участка меньше площади всего контура. Этот вид экстраполяции применяется для дешифрирования и оконтуривания отдельных ПТК.
Внутриландшафтная экстраполяция
осуществляется путем переноса дешифровочных признаков с изученного контура на контуры однотипных природных комплексов. Для этого выбирается наиболее характерный рисунок какого-либо объекта, в его пределах закладывается ключевой участок и разрабатываются дешифровочные признаки. Ареал экстраполяции ограничивается границами одного ландшафта.
Межландшафтная экстраполяция
осуществляется путем выявления однотип-
ных контуров в ландшафтах-аналогах. Ареал экстраполяции связан с рубежами физи- ко-географического районирования. Достоверность экстраполяции тем выше, чем ближе в классификационной системе и в сетке районирования лежат ландшафты-аналоги.
7.3. Ботаническое содержание биогеографических карт
Среди биогеографических карт особое место занимают геоботанические карты, отображающие не только растительность как таковую, но и ее связи с окружающими условиями и с животным миром (Соча- ва,1979). Отечественная школа геоботани- ческого картографирования, основы которой заложены Н. И. Кузнецовым, Е. М. Лавренко, В. Б. Сочавой занимает лидирующее положение в мире.
Картографирование зоологических объектов намного сложнее. На помощь приходит ландшафтно-экологический подход, когда в качестве основных объектов картографирования выступают природно-терри- ториальные комплексы и растительные сообщества, с которыми связаны местообитания определенных животных.
В разделе, посвященном топологическому уровню биогеографических исследований (см. гл. 6), отмечалась связь пространственной структуры растительного покрова с характером экотопических условий, с морфологической структурой ландшафта. Можно с уверенностью сказать, что каждому типу ПТК отвечает своя совокупность растительных сообществ. Более того, специфика ПТК определяет особенности их хозяйственного использования и соответствующий отклик экосистем на антропогенные воздействия. При определенной полноте экологических сведений о связи животных с ПТК и растительностью зоогеографические карты составляются методами индикации и экстраполяции (Вернандер и др., 1959).
При крупном масштабе аэрокосмических изображений на основе отдешифрированных контуров фаций и урочищ, ассоци-
111
аций и групп ассоциаций обнаруживаются семейные участки, строение и взаиморасположение убежищ, топография участков ареала; при среднем масштабе снимков и карт – на основе типов урочищ, их сочетаний, групп и классов ассоциаций, формаций выявляется топография частей ареала; на обзорных картах – на основе урочищ доминантов, ландшафтов, групп формаций, типов растительности – топография крупных частей ареала (Тупикова, Комарова, 1979).
Остановимся более подробно на геоботаническом содержании карт. Объектом картографирования является все разнообразие растительных сообществ – естественных, естественно-антропогенных, синантропных сообществ сорной растительности, агроценозов, динамические серии или разные стадии восстановления коренной растительности и т. д. Особенность отображения растительного покрова состоит в том, что растительность может картироваться не сама по себе, а как элемент ландшафтной структуры: растительность фаций, урочищ и т. п. или как элемент природнохозяйственных территориальных систем: растительность сельскохозяйственного контура, городского парка и др.
Легенда карты включает или классификационные таксоны сообществ (ассоциации, группы ассоциаций, формации и т. п.), или звеньев ординационных рядов (ряды сообществ, связанные осью изменения тех или иных экологических факторов), или комбинации сообществ (микрокомбинации сообществ фаций, мезокомбинации сообществ урочищ и т. п.), или динамические серии, или ряды трансформации синантропных растительных сообществ.
Чем крупнее масштаб карты, тем больше возможностей показать на карте низшие единицы классификации растительности – группы ассоциаций или даже ассоциации.
Закономерности распределения фитоценозов, соотношения между характером растительного покрова и условиями обитания познаются с помощью экотопологических профилей. Метод профилей позволяет представить конкретные сообщества в их
пространственной сопряженности друг с другом и меняющимися условиями среды. Для документального обоснования работы требуется наличие конкретных геоботани- ческих описаний с указанием места на карте. Составление по возможности детального списка растительных ассоциаций исследуемой территории – одна из важнейших задач полевых исследований. Эти данные представляют большую ценность для выявления пространственно-временных смен растительности.
На основе закономерностей, выявленных с помощью экотопологических профилей, составляются обобщенные экологические ряды растительности, т. е. некоторые абстрактные схемы, отражающие характерную для данного ландшафта смену фитоценозов под влиянием направленного изменения интенсивности какого-либо одного комплекса экологических факторов: режима увлажнения, богатства или засоленности почв и т. п.
Каждый ландшафт характеризуется своим спектром экологических рядов. Удобной формой, раскрывающей порядок экотопологических смен, являются экологические кресты, составляемые по методу В. Н. Сукачева. Крест строится в виде системы координат, центр которой занимают “средние” условия существования и типич- ные для них растительные сообщества. Направление осей указывает изменение того или иного экологического фактора и связанное с ним экологическое изменение растительности. Такие кресты наглядно отображают экологические ареалы ассоциаций, экологическую позицию одних сообществ по отношению к другим. Звенья экологи- ческих рядов могут включаться в легенду, экологический крест полезно давать на врезке к геоэкологической карте.
Континуальность и дискретность растительного покрова. Обсуждая методы показа растительного покрова на карте, мы должны коснуться вопроса о характере границ сообществ и явления континуума.
Наряду с дифференциацией растительного покрова наблюдаются процессы его интеграции. Когда границы между сообще-
112

ствами резкие, растительный покров предстает в виде дискретных единиц. Однако часто границы между фитоценозами предстают в виде постепенных переходов, и тогда возникает явление континуума, т. е. непрерывности растительного покрова.
Идея непрерывных переходов находит все большее признание как у биогеографов, так и у ландшафтоведов (Экотоны ..., 1997). Г.А. Исаченко отмечает, что дискретное видение геокомплексов как иерархических природных единиц с “жесткими” границами малопомалу сменяется континуальными подходами, более соответствующими глубинной сущности сложных природных систем (Исаченко,1994). Разграничение растительных сообществ осложняется многолетней динамикой ландшафтов, сложными механизмами взаи-
модействия природных процессов и антропогенных воздействий, несовпадением во времени и в пространстве изменений различных компонентов. В биосфере калейдоскоп сменяющих друг друга биоценозов в конечном счете образует пространственновременной континуум, в котором все сообщества связаны друг с другом сериями переходов.
В. С. Ипатов и Л. А. Кирикова (1985), детально рассмотрев причины проявления континуума и дискретности, пришли к выводу, что континуум и квантованность являются коренными свойствами растительного покрова и выражены одновременно и всюду. Определив континуум как свойство растительного покрова, выражающееся в том, что два любых произвольно выбранных, примыкающих друг к другу участка всегда имеют общие признаки, авторы под- черкивают, что это свойство проявляется как на уровне растительного покрова в целом, так и на уровне его ценоэлементов.
Явления дифференциации и интеграции растительного покрова наглядно изоб-
ражает Р. Уиттеккер (1980), рассматривающий несколько рабочих гипотез возможных вариантов распределения видовых популяций (рис. 7.4).
Конкурирующие виды, включая расте- ния-доминанты, взаимно исключают друг друга, и их популяции разделены резкими границами. В этом случае вдоль градиента возникают различающиеся между собой сообщества, отделенные друг от друга четкими границами (A). Конкурирующие виды-доминанты разделены резкими границами, однако множество сопутствующих видов образуют перекрывающие друг друга переходы (B). В центре сообщества господствует специфичный для него набор видов, однако к периферии усиливается краевой эффект, проявляющийся в форме экотона: сообщества постепенно переходят друг в друга, образуя континуумы (C). Резкие границы между видовыми популяциями отсутствуют. Центры и границы видовых популяций более или менее равномерно рассеяны вдоль градиента среды (D).
С континуальностью растительного покрова связано явление экотона. В своем первичном понимании (Clements, 1936) экотоны представляют собой контактные “микрозоны” между растительными сообще-
113
ствами (экосистемами). Они отличаются выраженным краевым эффектом - повышенной численностью организмов и усиленным влиянием сообществ на окружающую среду.
В. С. Залетаев (1989), определив особенности экологически дестабилизированной среды, отмечает двойственность ее свойств. С одной стороны, это возникновение дробной экологической дифференцированности и контрастности, а с другой - усиление экологической связанности разнородных природно-территориальных комплексов на основе существенно возросшего транспорта веществ и расселений организмов.
При всем разнообразии конкретных экосистем В. С. Залетаев предлагает различать два главных принципа организации биогеоценозов: 1) “жесткой” и 2) “плавающей” связи. Первый принцип предусматривает формирование такой структуры экосистемы, при которой образующие ее виды функционируют в узком диапазоне значе- ний экологических факторов; эдификаторы сообщества относятся к стенобионтным видам. Б. А. Юрцев (1988) называет их дифференцирующими. Благодаря им одни сообщества отличаются от других. Они образуют биологический каркас биогеоценоза, обеспечивающий ему устойчивость в стабильных природных условиях. Растительный покров, организованный по принципу “жестких” связей, характеризуется большей дискретностью, обусловленной четкой приуроченностью к определенным экотопам. дифференцирующих видов.
Принцип “плавающей” связи проявляется в том, что устойчивость системы обеспечивается возможностью настройки ее к тем или иным флуктуациям среды за счет эврибионтных видов, их перегруппировки, изменения ритмов активности и т. п. Б. А. Юрцев назвал такие виды интегрирующими, они как бы сшивают экотопы разного класса благодаря тому, что обладают широким экологическим ареалом. Организация по принципу “плавающих” связей характерна для структуры растительного покрова на переходных экотонных территори-
ях. В этих условиях характерными оказываются групповые адаптации близких видов к переменным параметрам внешней среды. При этом возникает система взаимных функциональных дополнений, взаимозаменяемость видов, выступающих в экосистеме как функциональная целостность.
Комплексность растительного покрова. Комплекcность, в ее более общем понимании (включая и мозаичную структуру растительного покрова внутри сообщества), зависит от чередования определенных условий среды и воздействия человека. Особенно ярко она выражена на верховых болотах, в полупустыне и пустыне, на месте лесных вырубок, пастбищ, пахотных угодий и т. п.
Детальные исследования позволяют выявить неоднородность и лесной, и луговой, и степной растительности; и даже в моновидных агроценозах (например, посевах пшеницы), раскинувшихся на больших площадях, проявляется неоднородность растительного покрова, обусловленная неоднородностью рельефа и литогенной основы.
Комплексность создает определенные трудности в показе структуры растительного покрова на картах. Очень редко участок земной поверхности, занятый одной ассоциацией, оказывается достаточно большим, чтобы его можно было изобразить в виде отдельного контура.
Назовем типичные случаи неоднородности растительного покрова, которые можно вводить в легенду крупномасштабной карты (по В. В. Мазингу, 1969):
1.Одна пространственно господствующая единица содержит фрагменты других сообществ или окаймлена полосой резко различных типов сообществ, показать которые в данном масштабе не удается. Например, ирригационная сеть с деревьями и кустарниками вдоль каналов.
2.Поясное распределение сообществ, каждое из которых не может быть показано на карте. Например, берега небольших зарастающих озер, растительность западин
èò. ï.
3.Закономерные сочетания, состоящие из чередующихся местообитаний с харак-
114

терными сообществами или их фрагментами, которые в отдельности картировать невозможно. Например, комплексная растительность бугристых и грядовых песков.
Следует иметь в виду, что пренебрежение комплексными единицами приводит к грубому упрощению. Без учета комплексности растительного покрова геоботанические карты теряют свою экологическую определенность, страдает их конкретность.
Динамика растительности. Растительный покров – явление разновозрастное. Он прошел длительный путь развития вместе с географической оболочкой. Эволюционный путь развития необратим и характеризуется увеличением числа составляющих растительные сообщества элементов и усложнением их структуры. В отличие от этих эволюционных изменений растительности свойственны динамические превращения, обусловленные экологическими факторами или антропогенным воздействием.
В. Б. Сочава и его последователи предлагают выделять коренные и производные динамические категории, составляющие основу геоботанического картографирования.
Коренные сообщества. Им соответствует понятие зонального типа растительности. Они разделяются на абсолютно, практически и условно коренные.
Абсолютно коренные сообщества - это те, что сохранились еще от доагрикультурного и допромышленного развития. По-видимому, они очень редки на земном шаре. Практически коренные сообщества - это такие, в которых под влиянием антропогенных факторов незна- чительно изменились состав и структура, или такие, которые прошли полный ряд трансформаций до восстановления коренного растительного покрова. Условно коренные сообщества представляют собой более или менее устойчивые стадии в процессе развития нарушенного растительного покрова. Это, например, таежные леса, которые подвергались в прошлом пожарам и рубкам, а в настоящее время испытывают их, но в
ослабленной форме; при этом они во флористическом и экологическом отношениях сохраняют близость к исходным коренным лесам.
Динамические серии естественной растительности наблюдаются в ландшафтах с быстрым и направленным изменением природных условий. Например, в поймах крупных рек можно наблюдать последовательные смены растительности от пионерных стадий зарастания пляжей до вполне сформировавшихся климаксовых сообществ высокой поймы.
Производные сообщества. В ландшафтах, подверженных антропогенному воздействию, на месте коренной растительности формируются элементы техногенной инфраструктуры, сельскохозяйственные угодья и производные сообщества (рис. 7.5).
Длительнопроизводные растительные группировки возникают при систематическом антропогенном воздействии: рубках, пожарах, сенокосах и выпасах скота, пахоте и т. п. При этом наряду с изменениями в растительном покрове существенно изменяются почвообразование, ход геоморфологических процессов, гидрологический режим. Такие группировки зачастую относятся к иному типу растительности, чем коренные сообщества. В частности, в таежной зоне на месте коренных лесов возникают вторичные луга, болота, пустоши.
Понятие кратковременнопроизводные
сообщества относительно, ибо время восстановления коренной растительности зависит от ее типа и глубины антропогенного воздействия. Ряды трансформации синантропных растительных сообществ отражают сукцессии растительности в местах, где естественный покров был полностью уничтожен человеком. Например, стадии дернового процесса на пашне.
С прекращением антропогенного воздействия производные растительные сообщества путем ряда смен идут к восстановлению коренной растительности. Важно знать, что последовательность смен зависит от характера коренной растительности и всего комплекса природных факторов. Каждой коренной группировке, сформировавшейся на месте определенного природ- но-территориального комплекса, присущи свои ряды производных сообществ.
Понятие коренной растительности Р. Тюксен дополнил понятием потенциальной растительности, т.е. той, которая должна появиться в данных природных условиях вследствие ряда трансформаций производных сообществ после того, как вмешательство человека прекратится. Поскольку в результате антропогенного воздействия происходит глубокое нарушение не только состава и структуры растительных сообществ, но и всего комплекса условий обитания, восстановленная растительность, как правило, будет отличаться от коренной. Так, например, на смену уничтоженным человеком тропическим лесам приходят саванны.
7.4. Региональные аспекты биогеографических исследований
В современной биогеографии важнейшим направлением подготовки специалистов, а также научных и прикладных исследований признается рациональное природопользование и охрана природы в конкретных регионах.
Современная стратегия региональной экологической политики основывается на представлениях о природно-территориаль- ных комплексах (ПТК) как сложно организованных территориальных системах – геосистемах, экосистемах – разной размерности. Своим многообразием они обеспечи- вают устойчивость процессов саморазвития всей биосферы. Охрану растительности и животного мира следует решать с учетом их неразрывной связи с природными ландшафтами. Поэтому приоритетную роль в решении задач рационального природопользования и охраны природы должны играть ландшафтно-экологические исследования. Предыдущие разделы можно рассматривать как теоретическое обоснование данного утверждения. Достоверность теории подтверждается опытом. Приведем несколько примеров.
Процессы опустынивания и механизмы естественной стабилизации ландшафтов Северо-Западного Прикаспия. Прикаспийская низменность – единственная область в Европе, где степная зона сменяется зоной полупустыни; последняя широкой полосой продолжается к востоку, образуя северное обрамление азиатских пустынь. В России именно здесь наиболее ярко проявляются признаки опустынивания и экологической дестабилизации природных ландшафтов в результате антропогенного воздействия. Аридность и безводность Прикаспийской низменности, засоленность материнских пород и почв, низкая продуктивность растительности всегда препятствовали хозяйственному освоению этой обширной территории. Издавна она представляла собой область кочевого отгон-
116

ного скотоводства. Животноводческий профиль сохранился в Прикаспии и в настоящее время. Нерегулируемая пастьба скота, перевыпас являются основной причиной пасторальной дигрессии и деградации почвенного покрова, образования массивов подвижных песков.
Строительство оросительных и обводнительных систем на базе волжских вод способствовало развитию в Северо-Западном Прикаспии орошаемого земледелия. Однако потери воды в результате инфильтрации из необлицованных русел каналов, избыточное орошение являются при- чиной вторичного засоления почв, что делает их малопригодными для дальнейшего хозяйственного использования.
Недра Прикаспийской низменности весьма перспективны в нефтегазоносном отношении: в Астраханской области, в Казахстане уже открыты
уникальные по запасам месторождения нефти и газа. Интенсивные геолого-разведоч- ные работы ведутся на территории СевероЗападного Прикаспия. Они сопровождаются мощным воздействием гусеничного и колесного транспорта на растительный и по- чвенный покров полупустыни. В результате образуются техногенные формы рельефа, массивы подвижных песков. Некоторые скважины дают приток высоко минерализованных грунтовых вод, вызывая засоление прилежащих территорий.
Дельта Волги и прибрежная полоса Каспийского моря известны как места нереста и обитания разнообразных рыб, в том числе осетровых. В зарослях тростника гнездятся и находят убежище многочисленные птицы. Вместе с тем плоский рельеф побережья Северного Каспия является при- чиной того, что регрессивные или трансгрессивные фазы колебания уровня Каспийского моря оказывают глубокое воздействие на прибрежные экосистемы.
Усиливающиеся процессы экологической дестабилизации и опустынивания земель Северо-Западного Прикаспия делают
актуальной задачу изучения закономерностей этих процессов, чтобы на их основе выявить механизмы естественной стабилизации экосистем и восстановления их природного потенциала (Петров, 1996).
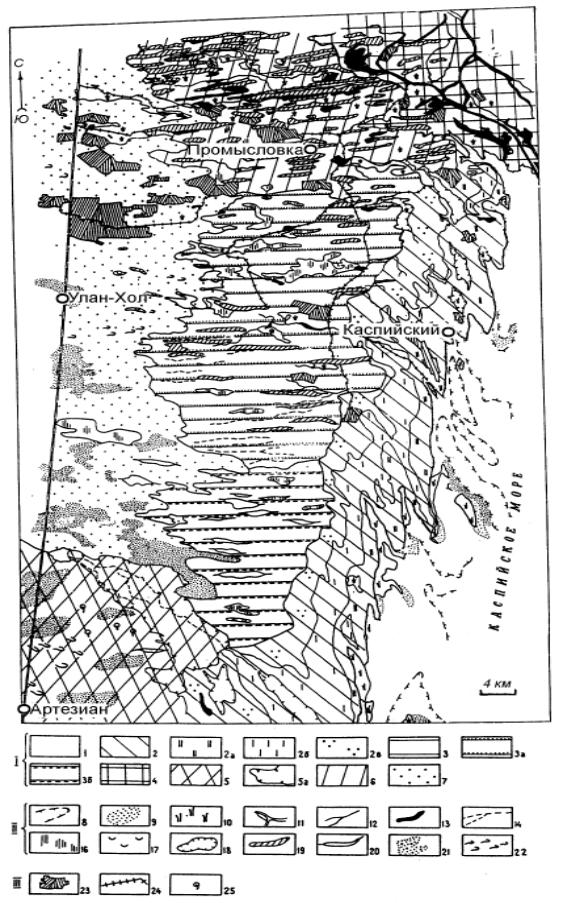
Морфологическая структура ландшафтов. По схеме физико-географическо- го районирования (Доскач, 1979) описываемая территория относится к области западного правобережного Прикаспия, к районам Центрально-Черноземельской равнины, Приморской песчано-солончаковой низменной равнины; с севера к ним примыкает район бэровских бугров и подстепных ильменей. Своеобразие каждого из названных районов хорошо видно на космическом снимке (рис. 7.6).
Полевые ландшафтно-экологические исследования, проведенные с использованием материалов дистанционного зондирования, позволили выявить детали морфологической структуры ландшафтов (рис. 7.7). На рис. 7.8 приведены аэрофотографические эталоны характерных ПТК.
Остановимся на краткой характеристике типичного для Центрально-Чернозе- мельского района ландшафта верхнехва-
117
118

Рис. 7.8. Аэрофотографические эталоны некоторых природно-территориальных комплексов Северо-За- падного Прикаспия. Масштаб 1 : 10 000:
а – характерный для Центрально-Черноземельского района ландшафт верхнехвалынской пологоволнистой равнины. Территория вдоль железной дороги к северу от пос. Улан-Хол. Видны прямоугольные контуры пахотных угодий, частично заброшенных, частично возделываемых; б – житняковопрутняково - полынная растительность пологоволнистой равнины. Наземная фотография; в – “степные блюдца”, обусловливающие мелкопятнистый рисунок верхнехвалынской равнины; г – бэровские бугры (светлые), разделенные подстепными ильменями (темные)
Рис.7.7. Урочища-доминанты Северо-Западного Прикаспия по материалам дешифрирования спектрозональных КС со спутника серии “Космос”:
Ландшафты: 1 – морские мелководья с зарослями прибрежно-водной растительности и песчано-ракушечными отмелями; 2 – современное низменное побережье Каспийского моря, первая приморская терраса от -25,5 до -29 м; 2а – нижний уровень приморской террасы от -27 до -29 м с тростниковыми плавнями на примитивных луговоболотных почвах; 26 – средний уровень приморской террасы от -26,7 до -27 м с кермеково-бескильницевыми сообществами на аллювиально-маршевых сильно солончаковых почвах; 2в – верхний уровень приморской террасы от -26,5 до -26,7 м с сообществами солончаковой полыни и однолетних солянок на солончаковых и солонча- коватых почвах; 3 – новокаспийская равнина, вторая приморская терраса от -22 до -25,5 м; За – плосковолнистая равнина с останцами бэровских бугров и лиманами; 3á – плоская равнина с сообществами солончаковой полыни, песчаными массивами, солончаковыми понижениями, сухими руслами, сглаженными формами бэровских бугров и лиманами; 4 – дельта Волги; 5 – верхнехвалынская песчаная бугристая равнина с житняково-прутняко- во-полынной растительностью на закрепленных песках и зарослями псаммофитов на эоловых массивах; 6 – ландшафт бэровских бугров и подстепных ильменей; 7 – верхнехвалынская супесчаная пологоволнистая равнина с житняково-прутняково-полынной растительностью. Урочища-доминанты: 8 – ложбины и впадины подводного берегового склона глубиной более 1 м; 9 – песчано-ракушечные отмели, лишенные растительности, глубиной 0,5 м;10 – песчано-ракушечные отмели с зарослями прибрежно-водной растительности глубиной 0,1– 0,25 м: 11 – рукава, протоки дельты Волги в сочетании с мелкими култучными озерами;12 – ерики, 13 – ильмени; 14 – сухие русла;15 – подтопляемые участки подстепных ильменей с гидрофитно-галофитной растительностью (тростник, солерос, шведка), 16 – солончаки,17 – западины с галофитной растительностью, сгруппированные но руслообразным ложбинам; 18 – солончаковые депрессии; 19 – бэровские бугры; 20 – останцы бэровскнх бугров с обарханенными вершинами; 21 – массивы эоловых песков; 22 – барханы. Антропогенные элементы: 23 – пашни: 24 – каналы; 25 – колодцы
119

Рис. 7.9. Элементы морфологической структуры и растительного покрова характерного для Центрально- Черноземельского района ландшафта верхнехвалынской пологоволнистой равнины:
I – II на рис. – фрагменты региональных профилей. Морфологические ПТК: ПСР – урочище супесчаной пологоволнистой равнины; Б.б – бэровский бугор, Рсл – сухое русло; МСО – солончаковая депрессия; ПМ
– песчаный массив; НР – новокаспийская равнина. Цифры внизу – номера точек наблюдения. Раститель-
ность: 1 – тамарикс; 2 – биюргун; 3 – прутняк; 4–6 – полыни: 4 – солончаковая, 5 – таврическая, |
6 |
– |
белая; 7 – сарсазан; 8 – кермек; 9 – верблюжья колючка; 10 – тростник; 11 – костер кровельный; |
12 – |
|
юзарлык; 13 – горяниновия; 14 – курай; 15 – верблюдка; 16 – однолетние солянки; 17 – эбелек; |
18 |
– |
житняк; 19 – джузгун; 20 – кумарчик; 21 – лебеда татарская; 22 – шведа |
|
|
лынской пологоволнистой равнины. Элементы его морфологической структуры и растительного покрова раскрывают фрагменты двух региональных ландшафтно-эко- логических профилей (рис. 7.9). Господствующая роль принадлежит урочищу-до- минанту слабоволнистой равнины с житня- ково-прутняково-полынной растительностью. На профиле данное урочище обозна- чено как ПСР. Кроме того, характерными элементами морфологической структуры являются: массивы заросших песков (ПП), подвижных песков (ПМ), межбугровых депрессий (МСО).
Растительный покров и его динамика. На плоской поверхности Прикаспийской низменности каждая неровность приводит к перераспределению скудной поверхностной влаги и значительным различиям в динамике всех процессов. Узор биогеоценотического покрова (фитоценохор) и сукцессии экосистем в значительной мере контролируются типами морфологи-
ческих комплексов, а также видами и интенсивностью хозяйственной деятельности. Не останавливаясь на сравнительной характеристике ПТК, отметим особенности растительности урочища-доминанта ПСР.
Верхнехвалынская пологоволнистая равнина интенсивно используется как зимнее пастбище. В результате перевыпаса и техногенного воздействия растительность деградирует и появляются массивы полузакрепленных или подвижных барханных песков (рис. 7.10).
Значительная часть территории вдоль железной дороги к северу от пос. Улан-Хол в обозримом прошлом была занята пахотными угодьями. В 70-е годы к этим землям были подведены оросительные каналы, и их вновь распахали. Засоленность почвогрунтов и грунтовых вод является причи- ной засоления орошаемых земель.
Вследствие интенсивной хозяйственной деятельности абсолютно коренная растительность не сохранилась. Она представ-
120