

Петров_К_М_Биогеография_СПб_2001
.pdfжие и сочные утром, блекли и свешивались |
доточились наиболее устойчивые, но бес- |
вниз, лепестки цветов осыпались. В июне |
плодные соединения алюминия и железа. |
и июле около полудня обыкновенно выпа- |
Продуктивность лесов огромна. Запас |
дает сильный дождь, приносящий желан- |
древесины свыше 5000 ц/га (в Бразилии – |
ную прохладу. Туча приближается всегда |
до 17 000 ц/га), а годовой прирост 350 – |
одинаковым, весьма любопытным манев- |
500 ц/га и более. Парадокс влажных тропи- |
ðîì. |
ческих лесов заключается в том, что пыш- |
Сначала прохладный морской бриз, по- |
ная растительность развивается на мало- |
являющийся около 10 час. утра и усилива- |
мощных бедных почвах. Это становится |
ющийся по мере того, как поднимается |
возможным благодаря очень быстрому кру- |
солнце, начинает ослабевать и, наконец, |
говороту биогенных солей. Отмершая мас- |
прекращается совершенно. Тогда жара и |
са ветвей и листьев быстро, в течение не- |
электрическое напряжение воздуха стано- |
скольких недель, разлагается и минерали- |
вятся просто невыносимыми. Тяжелое рас- |
зуется. В лесах мало опавшей листвы и не- |
слабление овладевает всеми, даже обитате- |
редко видны пятна оголенной земли. В раз- |
ли леса выдают это своими движениями. |
ложении опада большое участие принима- |
Но вот на востоке показывается белый ту- |
ют термиты; главная же роль, по-видимо- |
ман и начинает собираться в кучевые обла- |
му, принадлежит грибам. |
ка, нижний край которых становится все |
Корневая система деревьев не прони- |
темнее и темнее. Весь восточный горизонт |
кает на большую глубину, вся она сосредо- |
внезапно чернеет; эта мгла распространя- |
точена в первых десятках сантиметров ма- |
ется все большой и больше, заслоняя под |
ломощного слоя почвы, где образует фильтр, |
конец и солнце. Затем мощный порыв вет- |
поглощающий растворимые питательные |
ра с туманом проносится по лесу, раскачи- |
вещества. Важную роль в питании растений |
вая вершины деревьев, сверкает ослепи- |
играет микориза. Высокие деревья с повер- |
тельная молния, раздается удар грома и раз- |
хностной корневой системой мало устойчи- |
ражается стремительный ливень. Такие гро- |
вы. Досковидные корни – специальное при- |
зы быстро проходят и оставляют за собой |
способление, подобно контрфорсам у готи- |
неподвижные исчерна-синие тучи, остаю- |
ческих соборов они поддерживают высокие |
щиеся на небе до самой ночи. Вместе с тем |
стволы (рис. 9.5). |
вся природа освежена; лишь лепестки цве- |
Особенности почв влажных тропичес- |
тов и листья кучами валяются под деревья- |
ких лесов следует учитывать при их хозяй- |
ми. К вечеру природа пробуждается к но- |
ственном освоении. Издавна здесь приме- |
вой жизни, которой полон каждый куст, каж- |
нялась подсечно-огневая система земледе- |
дое дерево. На другой день солнце снова по- |
лия: лес вырубался, сжигался, участок об- |
является на безоблачном небе, возобновляя, |
рабатывался для выращивания тех или |
таким образом, круговорот тропического |
иных сельскохозяйственных культур (рис. |
дня, в течение которого перед нашими гла- |
9.6). Но запасы питательных веществ в по- |
зами как будто проходят весна, лето и |
чве ограничены, и они быстро вымывают- |
осень…” |
ся дождями. Через пару лет возделанный |
Почвы влажных тропических лесов |
участок приходится забрасывать. Губитель- |
формируются на мощной бесплодной фер- |
на сплошная вырубка лесов, особенно на |
раллитной (Al2O3, Fe2O3) коре выветрива- |
склонах, которая ведет к смыву почв и ин- |
ния. Основной запас питательных веществ |
тенсивной эрозии. Около 100 000 км2 òðî- |
сосредоточен в живых растениях. Полага- |
пических лесов подвергается деградации |
ют, что растения получили этот запас мил- |
ежегодно в результате антропогенного воз- |
лионы лет назад в процессе биохимическо- |
действия. Обнажение бесплодной феррал- |
го разложения материнских горных пород |
литной коры выветривания на больших |
богатого минералогического состава. В ко- |
площадях – угроза того, что влажные тро- |
нечном счете, в коре выветривания сосре- |
пические леса могут утратить способность |
151

Рис. 9.5. Досковидные корни, придающие большую устойчивость высоким стволам деревьев с поверхностной корневой системой, по А. Кернеру
152

Рис. 9.6. Выжигание тропического леса под сельскохозяйственное угодье и его последствия:
Справа – эрозия склонов после уничтожения лесной растительности
êсамовосстановлению и на их месте возникнут вторичные растительные сообщества с более бедным видовым составом и низкой продуктивностью.
Богатство биома влажных тропических лесов находит выражение в огромном разнообразии видов, жизненных форм и в сложной структуре лесного сообщества. 70% видового разнообразия растений составляют деревья (цветковые). Участие трав незначительно, обычно они растут на опушках леса. Преобладают травы, своими размерами не уступающие деревьям, бананы достигают высоты 6 м. Их подземные органы живут неопределенно долго.
Число видов деревьев на 1 га может превышать 100; переходя от одного дерева
êдругому, исследователь каждый раз обнаруживает новый вид. Лесные сообщества не отделены друг от друга четкими границами, а связаны постепенными переходами. Такое явление называется горизонтальной континуальностью растительного покрова.
Так же сложен вопрос о вертикальном сложении леса: между 30 – 40-метровыми гигантами, образующими верхний полог, и деревьями в 3 – 5 м существуют виды деревьев, образующие кулисообразные полога, перекрывающие друг друга (рис. 9.7). Это дает основание говорить о вертикальной континуальности, которая усиливается благодаря массе внеярусных растений – лиан и эпифитов.
Число видов эпифитов и лиан в дватри раза превосходит число видов деревьев (рис. 9.8). Особую жизненную форму
представляют деревья-душители, большинство из которых относится к роду Ficus. Благодаря птицам их семена попадают на ветви крон деревьев верхнего яруса. На первой стадии развития росток ведет жизнь эпифита. Однако сразу начинается быстрый рост корней: один корень устремляется в землю и снабжает растение водой и питательными солями; ответвления воздушных корней охватывают ствол хозяина и душат его (рис. 9.9).
Влажный тропический лес создает своеобразный микроклимат под своим пологом. Кроны деревьев смыкаются; в лесу ца-
Рис. 9.7. Вертикальная структура дождевого тропи- ческого леса, по Г. Вальтеру:
Разные виды деревьев высотой от 4 – 5 до 30 – 35 м образуют переходы, затрудняющие выделение четких элементов вертикальной структуры лесного сообщества
153

рит зеленый сумрак, редкий луч солнца пробивается сквозь густую листву. Здесь влажно и душно, как в оранжерее. Условия обитания накладывают отпечаток на строение растений. Стволы гладкие и стройные; кора тонкая и светлая. Корка, как правило, не образуется. У многих деревьев досковидные и ходульные корни. Почки защищены очень слабо, почечные чешуи отсутствуют. Форма и размер листьев в нижних ярусах леса мало варьируют: обычно они большие и цельнокрайние. В более высоких ярусах леса листья вырабатывают приспособления для защиты от яркого солнца и потери влаги – их размеры уменьшаются, они становятся кожистыми, блестящими, с толстой кутикулой. Представление о “вечнозелености тропических лесов” создается потому, что смена листьев в течение года происходит постепенно: одни листья разворачиваются
Ðèñ. 9.8. Орхидеи-эпифиты, по А. Кернеру из почек, другие опадают. Наряду с этим у некоторых деревьев может происходить одновременное массовое сбрасывание листвы, хотя рядом стоящие деревья того же вида могут оставаться зелеными.
В нижнем ярусе влажность воздуха постоянно высокая. Защиты от испарения растениям здесь не требуется. Напротив, у них есть ряд приспособлений, которые облег- чают транспирацию: тонкие листья с большой поверхностью и большим количеством устьиц; листья, в которых отдача влаги обеспечивается капельными устьицами на концах листовых жилок, из которых выдавливаются капли воды.
Многие тропические деревья цветут и плодоносят круглый год. На одном дереве имеются цветочные почки и цветки, зеленые и спелые плоды. Иногда деревья зацветают дружно раз в 10 лет. Учи- тывая, что деревья одного
вида в тропическом лесу растут на большом |
двудольных деревьев нужно назвать цейбу |
удалении друг от друга, приходится только |
(Ceiba) – дерево, возвышающееся над всем |
догадываться, благодаря какому сигналу они |
лесным пологом; бертоллецию (Bertolletia |
зацветают одновременно, обеспечивая тем |
excelsa), плоды которой называют бразиль- |
самым возможность перекрестного опыле- |
скими, или американскими, орехами; гевею |
ния. Одной из особенностей плодоноше- |
(Hevea brasiliensis), дающую каучук; деревья |
ния у некоторых видов растений является |
с ценной красной древесиной – махагонию |
каулифлория, т. е. образование цветков, а |
и цезальпинию. К нижнему ярусу относит- |
затем и плодов непосредственно на ство- |
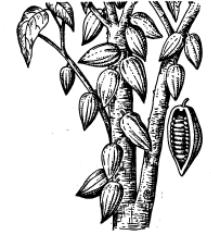
ся шоколадное дерево (Theobroma cacao); |
лах или толстых ветвях. Наиболее извест- |
дынное дерево (Carica, около 40 видов), |
ный пример – шоколадное дерево(рис. 9.10). |
сладкие плоды которого содержат фермент |
|
папаин, благоприятствующий пищеваре- |
|
нию. Деревья всех ярусов переплетают ли- |
|
аны. Среди эпифитов выделяются своим |
|
разнообразием орхидеи, папоротники, виды |
|
из семейства Bromeliaceae. В заводях Ама- |
|
зонки растет знаменитая гигантская кув- |
|
шинка виктория регия (рис. 9.13). |
|
Кроны густого тропического леса яв- |
|
ляются средой обитания множества зверей |
|
и птиц с ярким оперением (рис. 9.14). По- |
|
чти все они проводят большую часть жиз- |
|
ни на деревьях, не опускаясь на землю. |
|
Приспособление характерное для животных |
Рис. 9.10. Плоды шоколадного дерева (какао) на ство- |
неотропиков – цепкохвостость. Цепкие хво- |
ле и ветвях дерева. Рядом продольный разрез плода |
сты обезьян (рис. 9.15), опоссумов, древес- |
|
ных муравьедов и дикообразов, енотов-но- |
Животный мир влажных тропических |
сух помогают им ловко передвигаться с вет- |
лесов характеризуется огромным богат- |
ки на ветку. Типичными представителями |
ством и разнообразием: более 10 000 000 |
фауны Неотропического царства в гилее яв- |
видов, в основном насекомых и других бес- |
ляются неполнозубые – ленивцы. Их когти |
позвоночных (рис. 9.11). Благоприятные ус- |
срослись в серповидные крючья, с помощью |
ловия обитания – тепло, влажно, обилие зе- |
которых животные висят на ветвях безо вся- |
леного корма, масса убежищ, способствуют |
кого напряжения мускулов (рис. 9.16). |
формированию сообществ со сложными |
Впрочем, представление о том, что в |
биотическими отношениями. Круглый год |
тропическом лесу на каждом шагу встреча- |
климатические условия благоприятны для |
ются интересные птицы и звери, не соот- |
размножения и развития. Основная жизнь |
ветствует действительности. Девственный |
сосредоточена в кронах деревьев; у земли |
лес кажется мертвенно тихим: в густой чаще |
лес кажется безжизненным. |
трудно передвигаться и в то же время легко |
В Неотропическом царстве дождевые |
спрятаться. Лишь невидимые стаи обезьян |
тропические леса занимают огромные про- |
и птиц производят подчас невероятный |
странства на Амазонской низменности, где |
шум. Изредка встречаются равнинный та- |
их описал еще А. Гумбольдт, введший в на- |
пир и похожие на кабанов пекари. Крупные |
уку термин “гилея” (рис. 9.12). А. Л. Тахтад- |
грызуны замещают здесь копытных; наи- |
жян (1978) выделяет особую Амазон-скую |
больших размеров достигает капибара |
флористическую область, которая включает |
(Hydrochoerus capybara), до полутора мет- |
500 эндемичных родов и не менее 3000 ви- |
ров в длину и массой до 60 кг. Хищники |
дов. Здесь центр распространения пальм (до |
представлены кошачьими – ягуаром и оце- |
200 видов), из которых 60 эндемичны. Из |
лотом. |
155

Рис. 9.11. Мир насекомых обладает приспособлениями, делающими их трудно различимыми среди зелени листьев и цветов тропического леса
156

Рис. 9.12. Амазонская гилея, по А Кернеру:
На переднем плане монстера с воздушными корнями
157

Рис. 9.13. Виктория регия на Амазонке, по А. Кернеру
Важную роль в жизни влажных тропи- ческих лесов играют термиты – главные пожиратели отмершей растительной массы, благодаря чему она быстро разлагается и минерализуется. Термиты строят свои термитники, начиная с глубоких слоев почвы и кончая ветвями крупных деревьев. Основой сообщества является “царица” – крупная, до 12 см в длину, самка, непрерывно продуцирующая яйца (рис. 9.17). Количе- ство термитов на 1 м2 – от 500 до 10 тыс. Переваривание клетчатки в кишечном тракте термитов осуществляется с помощью одноклеточных жгутиковых, без симбиоза с которыми термиты существовать не могут. Поэтому появившуюся на свет личинку, рабочий термит подносит к анальному отверстию царицы, чтобы новорожденная обогатила флору своего кишечника.
Тропический лес от почвы до крон самых высоких деревьев заполняет множество видов муравьев. У ряда растений выработались особые симбиотические отношения
– мирмекофилия (любовь к муравьям): в стволах и ветвях деревьев возникают специальные пустоты, в которых живут хищные муравьи, уничтожающие насекомыхвредителей. Муравьи-листорезы ведут иной образ жизни. Они искусно вырезают кусочки листьев, переносят их в свои подземные гнезда, пережевывают и на этой разлагающейся массе выращивают опреде-
ленные виды грибов, которыми выкармливают личинок. Когда самка муравья-листо- реза отправляется в брачный полет, она берет в рот кусочки гифов гриба, что создать плантацию на новом месте.
Дождевые тропические леса, более подробно рассмотренные на примере Амазонской гилеи, сохраняют свои главные черты и в других областях своего распространения. Индивидуальный отпечаток накладывает их принадлежность к разным биогеографическим царствам.
В Эфиопском царстве дождевые тропические леса с мощным древесным ярусом, обилием лиан и эпифитов занимают большую часть бассейна Конго, а также побережье Гвинейского залива. В составе флоры 13 000 видов; более 250 родов растений эндемичных или преимущественно свойственных этим лесам. Широко представлены виды из тутовых, бобовых, пальмовых, стеркулиевых, панданусовых, и других семейств. В древесном ярусе господствуют рода: Ficus, Treculia, Chlorophora, Coula, Albizzia, Copaifera, Khaya, Ceiba, Cola, Omphalocarpum.
Обитатели тропических лесов (см. рис. 8.8) – гориллы, шимпанзе и различные мелкие обезьяны; из грызунов – длиннохвостая летяга; из копытных – малый оленек и окапи; из хищников – леопард. Из птиц следует упомянуть серого попугая, длинноклювого токо, птицу-носорога, цесарку и африканского павлина.
Характерным приспособлением к постоянному обитанию в кронах деревьев у животных является планирование. Способностью к планирующему полету с дерева на дерево обладают не только птицы, но и белки, змеи, ящерицы и лягушки.
В Индо-Малайском царстве дождевые тропические леса занимают преимущественно полуостров Малакку и острова Суматра, Ява, Борнео, Целебес, Новая Гвинея и соседние более мелкие острова.
Флора подобласти одна из наиболее богатых флор Земли (около 45 тыс. видов растений); эндемичных родов около 500; видовой эндемизм чрезвычайно велик. Так, например, на Новой Гвинее около 12% ро-
158

Рис. 9.14. Характерные представители фауны дождевого тропического леса. Бразильская область Неотропи- ческого царства. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – ленивец; 2 – средний муравьед; 3 – цепкохвостый дикобраз; 4 – попугай ара; 5 – тукан, 6 – игрунка;
7 – опоссум; 8 – большой муравьед; 9 – ревун; 10 – броненосец гигант, 11 – тапир; 12 – водосвинка
159

Рис. 9.14 (продолжение):
1 – пекари; 2 – ягуар; 3 – носуха; 4 – щелезуб; 5 – нутрия; 6 – колибри; 7 – паукообразная обезьяна; 8 – óäàâ áîà; 9 – бразильский попугай; 10 – гоацин; 11 – игуана; 12 – кракс, или шлемоносная курица
160