

Петров_К_М_Биогеография_СПб_2001
.pdfБиомы сухих субтропиков. Эти биомы характеризуются жарким и сухим летом, прохладной и дождливой зимой. Такой климат получил название средиземноморского, а биомы общее наименование – биомы средиземноморского типа (БСТ). Они располагаются в Северном и Южном полушариях между субтропическим поясом высокого атмосферного давления и областями зарождения циклонов в умеренном поясе – это Средиземноморье, Калифорния, Среднее Чили, Южная Африка, юг Австралии. Летом господствующим является воздействие центров высокого атмосферного давления, поэтому стоит жаркая и сухая погода; зимой в субтропики спускаются из умеренного пояса центры низкого давления, что сопровождается усилением циклонической деятельности, дождливой погодой.
ÂБСТ преобладают формации жестколистных (склерофилльных) вечноили зимнезеленых кустарников. Из многочисленных местных названий БСТ наиболее известны: маквис, гаррига, фригана и шибляк в Средиземноморье, чапарраль в Америке, финбош в Южной Африке и малли-скрэб в Австралии.
Происхождение вечнозеленых жестколистных кустарниковых формаций БСТ связывают с подлеском субгумидных лавролистных лесов. По мере усиления аридности климата древесный ярус этих лесов редуцировался, а кустарниковый подвергался изменениям в направлении увеличения числа склерофильных видов и их усиливающейся адаптации к сезонной сухости. В сухих лесах обычны низовые пожары, и адаптации к огню также могли начать формироваться уже под пологом леса.
Свидетельства о существовании типичных БСТ появляются не ранее позднего миоцена-плиоцена. Таким образом, БСТ представляются довольно молодыми биомами. В плейстоцене и голоцене ареал БСТ неоднократно то сокращался, то расширялся. Климатогенные пульсации в голоцене усугубились действием антропогенных факторов.
Âкачестве примера БСТ остановимся на биомах Средиземноморья. В начале тре-
тичного периода на равнинах господствовали тропические и субтропические леса, после поднятия гор сменявшиеся поясами широколиственных (платан, грецкий орех, каштан, тополь, дуб, бук и др.) и хвойных (сосна, ель пихта) лесов, отчасти местных, отчасти проникших с востока по гималайскому пути миграций. Межледниковые эпохи в плейстоцене вызвали формирование засушливых центров на Пиренейском полуострове, в Северной Африке и Малой Азии. Они обусловили ксерофитизацию и сильное обеднение когда-то богатой средиземноморской флоры. Тем не менее, именно флора третичного Средиземья явилась ядром, из которого развилась в основном на месте своего происхождения, т. е. автохтонно, современная средиземноморская флора.
Формирование современных БСТ происходило в условиях субтропического климата с жарким сухим летом и влажной зимой. Средние температуры зимы 7 – 14 îС, хотя абсолютные минимумы могут достигать -15 îС. Годовая сумма осадков 400 – 700 мм. Дефицит влаги (превышение испарения над выпадающими осадками) достигает 1000 – 1500 мм.
Берега Средиземного моря гористые. В нижней части склонов господствует вечнозеленая жестколистная (склерофильная) древесная и кустарниковая растительность на коричневых почвах. Характерный вид– маслина (Olea europea); ее ареал почти точно совпадает с ареалом БСТ (рис. 9.74, 9.75). Средиземноморские ландшафты издавна освоены человеком и несут яркие следы его деятельности – местами созидательной, местами разрушительной.
Господствующими формациями являются маквис, гаррига, фригана и шибляк.
Маквис – заросли с преобладанием небольших вечнозеленых деревьев и кустарников с мелкими жесткими листьями. Наиболее обычны здесь земляничное дерево (Arbutus unedo), мирт (Myrtus communis), ладанники (Cystus), фисташка (Pistacia lentiscus), дикая маслина (Olea europea var. oleaster), вечнозеленый каменый дуб (Quercus ilex) (рис. 9.76). Лианы – сассапариль (Smilax aspera), ломонос (Clematis
191

Рис. 9.74. Ареал маслины (Olea europea), по В. В. Алехину |
Рис. 9.75. Роща маслин |
|
flammula), плющ (Hedera helix) делают заросли трудно проходимыми. На юге Испании распространены заросли карликовой пальмы (Chamaerops humilis) – единственного вида пальм, имеющего свой естественный ареал в Европе.
Рис. 9.76. Маквис (Quercus ilex, Pistacia lenticus, Euphorbia dendroides, Rhamnus alaternus, Spartium juncus) на средиземноморском побережье Италии
Гаррига развивается на сухих каменистых почвах по южным склонам гор. Деревья отсутствуют, господствуют вечнозеленые кустарники с примесью дуба (Quercus coccifera), называемого на юге Франции “гарулия” (отсюда гаррига).
Фриганаразвивается в условиях сильной и продолжительной летней засухи, а также в местах усиленного выпаса как крайний вариант дигрессии растительного покрова.
Растения растут разреженно, всюду видна скелетная почва. Господствуют колю- чие, пахучие, часто сильно опушенные полукустарники и многолетники с одеревеневшими основаниями; весной обильно цветут эфемеры и эфемероиды. В Испании на гипсовых плато в районе Сарагосы фри-
гана представлена ассоциациями с господством солнцецвета (Helianthemum), розмарина (Rosmarinus officinalis), тимьяна (Thymus vulgaris), полыни белой (Artemisia herba-alba) с участием степных злаков: ковыля (Stipa parviflora), типчака (Festuca valesiaca) и др. (рис. 9.77).
Подобный ландшафт казалось бы не оставляет места для занятия земледелием. Однако на аэрофотоснимке этого района виден своеобразный членистый рисунок сухих русел, отображающий рукотворные терраски, удерживающие мелкозем и весеннюю влагу. Они используются для выращивания ячменя (рис. 9.78).
Шибляк представляет северный вариант средиземноморской растительности на черноморских склонах гор Крыма и Кавказа (рис. 9.79). Он образован зарослями листопадных форм деревьев и кустарников: дуба пушистого (Quercus pubescens), грабинника (Carpinus orientalis), держи-дерева (Paliurus
Рис. 9.77. Фригана на гипсовом плато в Испании
192

|
капливается преимущественно в сухой и |
|
|
жаркий сезон, когда дефицит влаги лими- |
|
|
тирует активность сапрофагов. Большин- |
|
|
ство почвенных беспозвоночных проводит |
|
|
лето в состоянии покоя или в фазе низкой |
|
|
численности. Таким образом, период наи- |
|
|
большей массы корма и период благопри- |
|
|
ятной погоды для потребителей не сов- |
|
|
падают. В результате биологическое разру- |
|
|
шение подстилки происходит медленно, за |
|
|
3 –5 лет. Существенным фактором, ограни- |
|
|
чивающим деятельность консументов, явля- |
|
|
ется жестколистность. Жесткие листья мало |
|
|
питательны, защищены механически и |
|
|
обычно слабо поедаются как фито-, так и |
|
|
сапрофагами. |
|
|
Из-за низкого уровня потребления пер- |
|
|
вичной продукции в БСТ образуется боль- |
|
|
шой запас сухой подстилки, на 70 – 80% со- |
|
|
стоящей из листового опада. Эта подстил- |
|
|
ка, препятствуя возобновлению, должна |
|
|
была бы делать мертво-покровные сообще- |
|
Рис. 9.78. Террасовое земледелие в Испании: |
ства БСТ неустойчивыми во времени. Это- |
|
го, однако, не происходит, поскольку БСТ |
||
а – фрагмент аэрофотоснимка, М. 1:5000. Виден членис- |
||
повсюду характеризуются высокой частотой |
||
тый рисунок террасированных сухих русел; б – земле- |
||
дельческие террасы в сухом русле, прорезающем повер- |
пожаров, которые в БСТ являются в значи- |
|
хность гипсового плато, занятого фриганой |
тельной мере природным, а не антропоген- |
|
|
ным феноменом. При снижении частоты |
|
spina-christi) и др. в сочетании со степны- |
пожаров они делаются более сильными и |
|
ми злаками и разнотравьем. |
разрушительными из-за увеличения запаса |
|
Животный мир Средиземноморья |
горючего материала. Значение пожаров для |
|
(рис. 9.80) сильно изменился под влияни- |
поддержания БСТ подтверждается тем, что |
|
ем многовековой деятельности человека. В |
их искусственное исключение в течение 60 |
|
настоящее время ведется большая работа по |
- 100 лет приводит к почти полной оста- |
|
его охране; единственный вид европейских |
новке возобновления доминантов. Истори- |
|
обезьян живет в естественных условиях в |
ческая типичность пожаров в БСТ, так же |
|
заповеднике на Гибралтарской скале. |
|
|
В БСТ растительная масса слабо ис- |
|
|
пользуется консументами как в пастбищных, |
|
|
так и в детритных пищевых цепях (Жери- |
|
|
хин, 1995). Эта особенность хорошо объяс- |
|
|
няется климатическим режимом. Для фито- |
|
|
фагов прямое климатическое ограничение |
|
|
связано с пиком прироста фитомассы в наи- |
|
|
более прохладный и дождливый период, |
|
|
когда неблагоприятная погода не позволяет |
|
|
растительноядным беспозвоночным не |
|
|
только достичь высокой численности, но и |
|
|
поддерживать постоянно высокую актив- |
|
|
ность. Растительный опад, напротив, на- |
Рис. 9.79. Шибляк на южном берегу Крыма |
|
|
193

Рис. 9.80. Характерные представители фауны Средиземноморской области Голарктического царства, по А. Г. Воронову:
1 – муфлон; 2 – протей; 3 – тосканская землеройка; 4 – белоголовый сип; 5 – канарейка; 6 – лань; 7 – гюрза; 8 – желтопуз; 9 – геккон; 10 – шакал; 11 – греческая черепаха; 12 – смирнский белый зимородок; 13 – пятнистая кукушка; 14 – дикобраз; 15 – франколин; 16 – дикий кролик; 17 – леопардовый полоз; 18 – горная курочка; 19 – генетта; 20 – бесхвостый макак
194
как в саваннах, подчеркивается широким распространением адаптаций к воздействию высоких температур у его обитателей.
Таким образом, в БСТ пожары являются постоянно действующим фактором и играют важнейшую роль в функционировании экосистем, обеспечивая возврат в оборот биогенов, связанных в мортмассе. По существу, огонь выполняет в этих экосистемах функцию своеобразного небиологического редуцента, действующего эпизодически, но чрезвычайно быстро, и придающего обороту системы своеобразную многолетнюю ритмику. Без пожаров устой- чивое существование БСТ немыслимо. Возврат биогенов в круговорот веществ опла- чивается ценой высоких потерь, особенно азота, в результате вымывания атмосферными осадками. Важнейший биологический компенсаторный механизм – широко распространенная в БСТ азотфиксация, к которой способны не только такие классические азотфиксаторы как бобовые, но и многие другие растения.
9.2. Биомы холодных и умеренных поясов
От полюсов к субтропическим широтам простираются холодные и умеренные пояса. Если в жарком поясе ведущим фактором формирования зональных типов биомов является возрастающая сухость климата, то в холодных и умеренных поясах главную роль играет первичный фактор географической зональности – температурный режим. Различия в климате и особенности палеогеографии накладывают свой отпечаток на своеобразие ландшафтов. Наряду с биполярностью органического мира отмеча- ются существенные черты отличия зональных типов биомов холодных и умеренных поясов Северного и Южного полушарий.
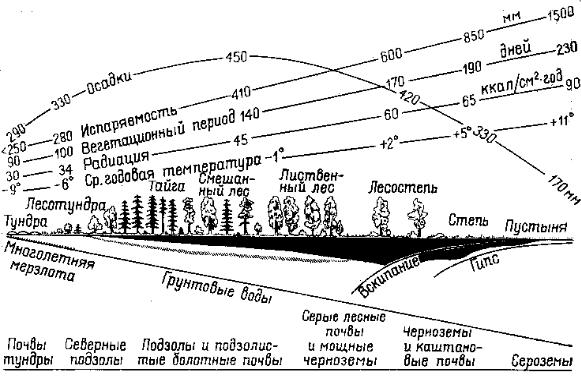
В Северном полушарии приполюсное пространство занято океаном, полярные пустыни представлены прерывисто в основном на островах. Биомы тундры и тайги имеют субширотное простирание, протягиваясь в виде концентрических полос через континенты Северной Америки и Евразии, принадлежащих к одному Голарктическому
Рис. 9.81. Схема изменения климата, растительности и почв на профиле Восточной Европы в направлении с северо-запада на юго-восток к Прикаспийской низменности, по А. П. Шенникову и Г. Вальтеру: Зачернен гумусовый горизонт; мощные черноземы соответствуют лесостепи. Заштрихован иллювиаль-
ный горизонт В
195

биогеографическому царству. Но по мере перехода в низкие широты, где климат становится более теплым, все отчетливее проявляется роль увлажнения, и в распределении зональных типов биомов находит выражение закон географической секторности (рис. 9.81).
В Южном полушарии приполюсное пространство занято Антарктическим континентом, лежащим в зоне полярных пустынь. Субантарктические и умеренные широты заняты в основном океанами. Наземные биомы формируются на островах и окраине Южной Америки. Типичные тундры, тайга и широколиственные леса, характерные для Голарктического царства Северного полушария, здесь отсутствуют. Зональные типы биомов Голантарктического царства обладают своими специфическими свойствами.
9.2.1. Тундра
Циркумполярное положение холодных поясов определяет суровость климата: отрицательный радиационный баланс, отрицательные средние годовые температуры воздуха. Границей холодного пояса служит изотерма самого теплого месяца 10 îС. Наряду с климатическими условиями большое влияние на формирование биомов холодных поясов оказывает взаиморасположение суши и океана.
Холодный пояс Северного полушария называют еще Арктическим или Арктикой. От полюса к югу тепловой режим становится более благоприятным для жизни. Это служит основанием для разделения Арктики на зоны и подзоны (рис. 9.82).
Тундры – главный зональный тип биомов холодного пояса Голарктического царства. Если весь холодный пояс называют Арктикой, то тундры с расположенной южнее лесотундрой относят к Субарктике.
Астрономической границей холодных поясов, где формируются биомы полярных пустынь и тундр, являются полярные круги – северный и
южный, располагающиеся на 66î 33‘ с. ш. и ю. ш. У полюсов солнце встает и садится лишь один раз в год. Полярный день и полярная ночь, начинающиеся и заканчивающиеся в дни равноденствий, длятся по шесть месяцев только на самих полюсах. У полярных кругов с нача- лом весеннего равноденствия продолжительность дня увеличивается ежедневно на 8 мин. Только одни сутки, в день летнего солнцестояния, солнце не заходит за горизонт; после осеннего равноденствия продолжительность дня быстро уменьшается.
Рис. 9.82. Зональное подразделение Арктики, по В. Д. Александровой:
1 – южная граница тундры, в том числе подзоны южной тундры;2 – южная граница подзоны северной тундры; 3 – южная граница зоны арктических пустынь
196

Тундра в пере- |
|
|
|
воде с финского |
|
|
|
языка означает |
|
|
|
плоское безлесное |
|
|
|
ï ð î ñ ò ð à í ñ ò â î . |
|
|
|
Ô î ð ì è ð î â à í è å |
|
|
|
тундровых биомов |
|
|
|
определяется суро- |
|
|
|
выми климатичес- |
|
|
|
кими условиями. |
|
|
|
Зима длинная, до 8 |
|
|
|
месяцев. Средняя |
|
|
|
годовая температу- |
|
|
|
ðà íèæå íóëÿ. Îä- |
|
|
|
нако не зимние хо- |
|
|
|
лода (полюс холо- |
|
|
|
да Северного по- |
|
|
|
лушария распола- |
|
|
|
гается, как извест- |
|
|
|
но, в Восточной |
|
|
|
Сибири в таежной |
|
|
|
зоне), а холодное и |
|
|
|
короткое лето опре- |
|
|
|
деляют особеннос- |
|
|
|
ти среды обитания |
|
|
|
в тундре. Замороз- |
|
|
|
ки возможны в лю- |
Рис. 9.83. Полярная граница древесной растительности, ограничивающая субарктичес- |
||
бой летний день, |
кую зону на юге, по Б. А. Тихомирову: |
|
|
но бывают и жар- |
Границу образуют: 1 – Betula tortuosa; 2 – Â. ermani; 3 – Picea obovata; 4 – P. mariana; |
||
кие дни, когда тем- |
5 – Larix sibirica; 6 – L. dahurica |
|
|
пература воздуха поднимается до 25 – 30 îÑ. |
ин Евразии. В Атлантическом секторе, где |
||
Атмосферных осадков (в основном в виде |
сказывается отепляющее влияние Гольфст- |
||
снега) выпадает немного 300 – 400 мм. Сне- |
рима, она поднимается выше Полярного |
||
говой покров маломощный, но он играет |
круга, в Тихоокеанском секторе под влияни- |
||
важную защитную роль, предохраняя нахо- |
ем холодных течений опускается к |
60î |
|
дящиеся под ним растения, а также живот- |
ñ. ø. |
|
|
ных от сильных морозов, ветра и поземки. |
В тундрах Голарктического царства еще |
||
В связи с низкими температурами испаряе- |
свежа память о недавнем оледенении: по- |
||
мость низкая, что приводит, несмотря на не- |
чти повсеместно распространены многолет- |
||
большое количество осадков, к избыточно- |
немерзлые породы (“вечная мерзлота”). Их |
||
му увлажнению: заболачиванию и образо- |
мощность изменяется от 300 – 400 м в арк- |
||
ванию многочисленных озер. |
тических тундрах, до первых метров в юж- |
||
От расположенной южнее зоны тайги |
ных тундрах. Глубина сезонного оттаивания |
||
тундру отделяет граница леса. Одним из |
колеблется от 30 – 40 см в северной полосе |
||
условий формирования таежной раститель- |
до 0,5 – 2 м в южной. Постоянно мерзлый |
||
ности является средняя месячная темпера- |
слой обусловливает охлаждение почвы и за- |
||
тура самого теплого месяца выше 10 îÑ (ðèñ. |
медление биохимических процессов, пре- |
||
9.83). |
|
пятствует фильтрации поверхностных вод |
|
Обращает внимание асимметрия гра- |
и способствует заболачиванию; он резко |
||
ниц тундры у западных и восточных окра- |
ограничивает грунтовое питание рек и их |
||
197

эрозионную деятельность. С мерзлотой связаны специфические геоморфологические процессы и формы мезо- и микрорельефа. Наиболее крупные формы – термокарстовые возникают в результате оседания грунта при вытаивании подземных льдов. Морозное растрескивание грунтов и образование ледяных клиньев в трещинах ведут к
Рис. 9.84. Полигональная тундра. Аэрофотоснимок
формированию полигонального микрорельефа (рис. 9.84). С мерзлотным пучением грунтов связан мелкобугристый рельеф. Криогенная сортировка твердого материала имеет следствием пятна-медальоны, структурные грунты.
Стабильность многолетнемерзлых грунтов, колебания верхней границы и даже само существование мерзлоты зависят от многих обстоятельств, и в первую очередь от растительного покрова, имеющего зна- чение важнейшего стабилизирующего фактора. Растительная дернина затрудняет теплообмен между мерзлым слоем и воздухом (поддерживает тепловое равновесие), скрепляет почву, препятствует сортировке грунта.
Недостаток тепла и избыток влаги, мерзлота, низкая емкость биологического круговорота, небольшое количество растительного опада, его замедленная гумификация и минерализация – все это определяет главные черты геохимических процессов и поч- вообразования. Химическое выветривание протекает слабо, высвобождающиеся основания вымываются из почвы, так что она обеднена кальцием, натрием, калием (но обогащена железом и алюминием). Широ-
кое распространение в Субарктике, особенно в южной ее части, имеют тундровые глеевые почвы с торфянисто-перегнойным горизонтом, под которым находится сравнительно однородная глеевая минеральная вязкая толща. В связи с низкой продуктивностью растительности торфообразование слабое; мощность торфа редко превышает 1 м. Почвенный покров, как и растительный, характеризуется мозаичностью.
Арктика – это нивальные ландшафты, мир снега и льда. Снежный покров играет важную роль в жизни растений и животных. Продолжительность залегания снежного покрова до 220 – 280 дней – главный отрицательный фактор в жизни большинства животных и растений. Толщина снежного покрова зависит от рельефа (с возвышений снег сдувается, а в понижениях накапливается), а также от континентальности климата – к западу от Таймыра она больше, к востоку меньше. В то же время снег играет и роль положительного экологического фактора. В качестве хорошего теплоизолятора он защищает почву, растения и животных от зимних морозов. Кроме того, слой снега предохраняет растения от снежной корразии: ветви, выступающие над снегом, срезаются или сильно повреждаются зимней поземкой, поэтому высота растений полностью зависит от мощности снегового покрова.
Тундровые биоты Палеарктики и Неарктики близки между собой. Многие виды растений: куропаточья трава (Dryas octopetala), брусника, водяника, альпийская толокнянка, альпийский мятлик и др., как и животных: северный олень, белый медведь, песец, белая сова и др. – имеют циркумполярный ареал. Общность тундровых биот можно объяснить общностью истории развития северных районов Евразии и Северной Америки, претерпевших оледенение. Биоты тундр характеризуются молодостью и бедностью видового состава: теплолюбивые формы третичного периода погибли или были коренным образом преобразованы во время четвертичного оледенения. Вместе с тем для Арктики характерен зна-
198

Рис. 9.85. Характерные представители фауны тундры. Рис. В. А. Ватагина, по Н. А. Бобринскому: |
|
|
1 – тундровая куропатка; 2 |
– белая куропатка; 3 – мохноногий канюк; 4 – лапландский подорожник; |
5 |
– пуночка; 6 – белая сова; 7 |
– белолобая казарка;8 – черная казарка; 9 – черношапочный сурок; |
10 |
– краснозобая казарка; 11 – копытный лемминг; 12 – обский лемминг; 13 – снежный баран; |
14 |
|
– северный олень; 15 – песец |
|
|
199

чительный процент эндемиков на родовом
èвидовом уровнях (рис. 9.85).
Âтундре отсутствует древесная растительность. По выражению М.В. Ломоносова: “Тундрами называют места мохом зарослые”. В растительном покрове кроме мхов и лишайников распространены низкорослые психрофитные
èкриофитные формы, приспособленные к существованию в условиях низких температур и высокой влажности почв, к использованию
тепла приземного слоя |
Рис. 9.86. Растения тундры, по Т. Н. Гордеевой и О. С. Стрелковой: |
|
|
и защитных функций |
À – крупка волосистая (Draba pilosa); Б – ива полярная (Salix polaris); В – карли- |
||
снежного покрова. |
ковая береза (Betula папа); Г – мак подушковидный (Papaver pulvinatum); |
Ä |
|
Растения тундр ха- |
|||
– дриада точечная, или куропаточья трава (Dryas punctata) |
|
||
рактеризуются низкой |
|
|
интенсивностью фотосинтеза, медленным |
роклимат, а нижние отмершие листья, по- |
|
накоплением органической массы, малой |
степенно разрушаясь, служат дополнением |
|
энергией роста. Годичные кольца растений |
к скудному содержанию азота в почве. |
|
нередко состоят из одного ряда клеток. По- |
У животных тундры также выработал- |
|
беги полярной ивы удлиняются за год на 1 |
ся целый ряд приспособлений, облегчаю- |
|
– 5 мм. Годичный прирост ягеля составляет |
щих существование в суровых условиях. |
|
около 4 мм. Однолетники в Арктике – яв- |
Например, у песца белая покровительствен- |
|
ление редкое; они не успевают завершить |
ная окраска (рис. 9.87.). Он быстро нагули- |
|
жизненный цикл в одно лето. Наиболее рас- |
вает толстый слой жира. Зимний мех его |
|
пространенная жизненная форма – карли- |
пушист и тепел – песец ценный пушной |
|
ковые кустарнички и многолетние травы |
промысловый зверь. Уши короткие (сравни- |
|
(рис. 9.86). Стебли у них короткие, листья |
те уши песца и пустынного зайца, |
ñì. |
малых размеров и у многих собраны в при- |
рис. 5.55). Подошвы густо покрыты мехом, |
|
корневую розетку. Обычны здесь и подуш- |
|
|
ковидные формы, возникающие при отмер- |
|
|
зании верхушечных почек. В таком случае трогаются в рост нижние пазушные почки, что приводит к обильному ветвлению и образованию коротких стелющихся побегов. Густо расположенные мелкие ветви с листьями на верхушках побегов придают кусту обтекаемую форму, которая хорошо противостоит ураганным ветрам. Под пологом
такой подушки создается более мягкий мик-
Рис. 9.87. Песец
200