

bkh_dlya_tebya
.pdfгруппами. Ксилулозу лучше запомнить относительно рибулозы у ксилулозы на 3-м углероде гидроксильная группа в другую сторону смотрит. Всего-то.
Моносахариды. Гексозы. Тоже 4 представителя: глюкоза, маноза, галактоза, фруктоза. Глюкоза, маноза и галактоза – альдозы, фруктоза – кетоза.
И опять-таки нет нужды запоминать все формулы – достаточно запомнить глюкозу и отличия от неё других гексоз. Глюкоза сверху альдегид, снизу CH2OH, дальше 4 гидроксильные группы – все смотрят вправо, кроме той, что на 3-м углероде. Маноза от глюкозы отличается тем, что у неё ещё и на 2-м углероде гидроксильная группа смотрит не туда. Галактоза отличается от глюкозы тем, что на её 4-м углероде гидроксильная группа смотрит не туда. Фруктоза отличается от глюкозы тем, что её 1-й и 2-й углероды поменялись карбонильной и гидроксильной группами. На рисунке, помимо линейных формул, приведены и циклические, которые вы просто обязаны знать. Запоминать циклические формы сложно – проще понять, как их выводить самостоятельно. На рисунке ниже на примере глюкозы я показал, как строятся циклические формы – вам это запоминать и писать не надо – вам надо просто понять. Значит есть молекула глюкозы, гидроксильная группа на 5-м углероде отдаёт свой водород кислороду альдегидной группы 1-го углерода, там образуется новая гидроксильная группа и свободная связь, с этой связью соединяется кислород бывшей гидроксильной группы 5- го углерода – образовался цикл. Теперь эту молекулу вы должны положить на бок, т.е. перевернуть на 90 градусов, тогда гидроксильные группы, что были слева, окажутся сверху, а те, что были справа, окажутся снизу. Рисовать нужно именно шестиугольник с кислородом – получается шестичленный цикл из соединительного кислорода и 5-ти углеродов. Теперь смотрим на формулу и решаем, как что расставлять в этом шестиугольнике. 1-й углерод –
131
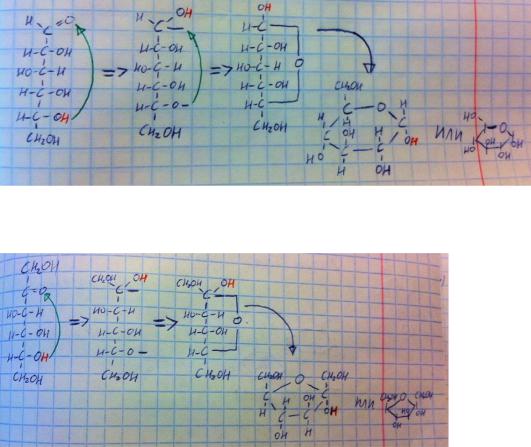
на нём раньше была карбонильная группа, но теперь гидроксильная группа, причём особо реактивная (она связана с углеродом, который связан с другим кислородом – остальные углероды в молекуле связаны только с одним кислородом), эта группа реагирует в химических реакциях в первую очередь,
а писать мы эту группу можем и вверх (бета-версия) и вниз (альфа-версия). 2- й углерод, его гидроксигруппа была вправо – теперь вниз. 3-й углерод, его гидроксигруппа была влево – теперь вверх. 4-й углерод, его гидроксигруппа была вправо – теперь вниз. 5-й углерод уже не имеет гидроксильной группы, ибо отдал её на образование цикла, зато у него есть CH2OH, который всегда торчит вверх. Вот и всё, это очень просто, если потренироваться. Советую научиться, ибо вам это не раз пригодится. В результате на рисунке в правом краю я дополнительно написал циклическую форму так, как её принято писать.
Это было правило для гексоз альдоз, а для фруктозы всё очень похоже, но немного отличается.
Здесь гидроксильная группа 5-го углерода взаимодействует с кетогруппой 2-го углерода, поэтому цикл пятичленный, логика расстановки гидроксильных групп та же. Первый углерод у нас CH2OH, гидроксильная группа на 2-м углероде, как и у глюкозы на 1-м, может смотреть и вниз (альфа) и вверх (бета), а первый углерод будет расположен на другом конце. Гидроксигруппа на 3-м углероде была слева – стала сверху. Гидроксигруппа на 4-м углероде была справа – стала снизу. 5-й углерод не имеет гидроксигруппу, зато CH2OH смотрит вверх. Ну вот и разобрались.
132
Что касается моносахаридов в нашем организме, то в подавляющем большинстве это пентозы и гексозы. Пентозы в основном – рибоза и дезоксирибоза – в составе нуклеотидов. Основной представитель гексоз – глюкоза, присутствует в составе разных соединений, в свободной форме находится только в крови. Основная фишка моносахаридов в нашем организме в том, что их нет в свободной форме нигде, исключение – глюкоза крови.
Теперь к олигосахаридам, как я и говорил, они имеют 2-10 мономеров
и в нашем организме в свободной форме не существуют, зато они существуют в составе гликопротеинов и гликолипидов. Гликолипиды (сфингогликолипиды) находятся в составе цитоплазматических мембран, особенно в нервной ткани, они образуют гликокаликс, выполняя защитную и структурную функцию, помимо этого они могут отвечать за транспорт веществ через мембрану. Гликопротеины (10% углеводного компонента, 90% белкового компонента) представлены практически всеми белками цитоплазматических мембран (защитная, транспортная ферментативная и многие другие функции, рецепторная), ферментами, рецепторами и некоторыми гормонами.
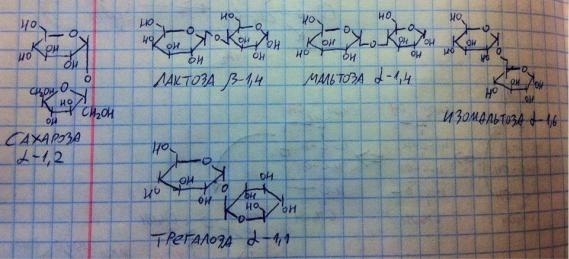
Мы должны ознакомиться с некоторыми представителями подкласса дисахаридов – их нет в нашем организме, но мы получаем их с пищей. 5
представителей: сахароза, мальтоза, изомальтоза, лактоза, трегалоза.
•Сахароза (тросниковый сахар, тот, что везде добавляют) состоит из глюкозы и фруктозы, связь альфа-1,2. Чем объясняется наименование связи? Связь образует гликозидный гидроксил, у глюкозы он (в данном случае) смотрит вниз, поэтому альфа, и расположен на первом атоме углерода, поэтому первая цифра – 1, а атом углерода, с которым связался гликозидный гидроксил, у фруктозы номер 2, отсюда двойка и альфа-1,2.
•Мальтоза (солодовый сахар, образуется при распаде крахмала) состоит из двух глюкоз, связанных альфа-1,4 связью.
133
•Изомальтоза образуется при распаде крахмала (подробности чуть позже) и состоит из двух глюкоз, связанных альфа-1,6 связью.
•Лактоза (молочный сахар) состоит из галактозы и глюкозы. Связь бета-1,4.
•И последний представитель, чью формулу рисовать необязательно, это трегалоза (содержится в грибах), состоит из двух глюкоз, связанных альфа- 1,1 связью.
Теперь наиболее широкий класс – полисахариды (имеют 10+ мономеров, обычно тысячи и десятки тысяч). Подразделяются на гомополимеры (состоят из одного типа мономера) и гетерополимеры (состоят из 2-х или более типов мономеров). У человека к гомополимерам относится гликоген – запасающий углевод, который кстати является разветвлённым, что помогает ему более компактно укладываться в клетках, мономером является глюкоза. К гетерополимерам углеводов относятся гликозамингликаны ГАГи, они строятся из дисахаридов, образованных гексуроновой кислотой и аминосахарами (те, в свою очередь, могут иметь и остатки кислот), получающиеся полимеры являются линейными и выполняют
в основном структурные функции, выделяют 6 классов ГАГ: гиалуроновая кислота, хондроитин-4-сульфат, хондроитин-6-сульфат, гепарин-сульфат, кератан-сульфат, дерматан-сульфат. А ещё существуют протеогликаны – комплекс ГАГ (90-95%) и белка (5-10%), белок из одной цепи, т.е. мономерный, функция протеогликанов тоже структурная. Протеогликаны и гликозамингликаны – неотъемлимые элементы межклеточного вещества всех соединительных тканей. Гликоген запасает много кто, но в первую очередь печень и скелетные мышцы.
Существуют сиаловые кислоты – производные глюкозы, входящие в состав некоторых веществ крови, содержатся в свободном виде в спиномозговой жидкости, входят в состав ганглиозидов (гликолипиды мембран нейронов) и участвуют в проведении нервного импульса.
Итак, мы закончили разбор классификации и представителей углеводов человеческого организма. Теперь можем приступать к самому обмену и начнём, как всегда, с переваривания.
Прежде, чем говорить о переваривании углеводов, надо разобраться, какие именно углеводы мы с пищей получаем – они разделяются на перевариваемые и неперевариваемые. К неперевариваемым относятся целлюлоза и пектин, которые ещё называют пищевыми волокнами, они имеют три очень важные функции: абсорбирование веществ, устройство среды для микрофлоры толстого кишечника и активация перистальтики. Чуть подробнее. Когда вы съедаете углеводы, они расщепляются в кишечнике в основном до глюкозы, которая в огромном количестве поступает в кровь, вызывая алиментарную гипергликемию, избыток поступающей глюкозы
134
превращают в жиры – это эффект быстрых углеводов. Если же в вашей пище присутствуют пищевые волокна, то большую часть глюкозы в кишечнике они абсорбируют, а потом отдают, но медленно – глюкоза поступает в кровь небольшими порциями, поэтому она может нормально утилизироваться без превращения её в жиры – медленные углеводы. К этому вопросу мы ещё вернёмся и всё станет понятнее. Что касается бактерий, в толстом кишечнике существует микрофлора, однако ей сложно закрепляться на гладкой и сколькой слизистой, а вот в пищевых волокнах им жить очень удобно – они прогрызают в них ходы для себя. Ну а с перистальтикой, думаю, всё понятно
– штука нужная. Теперь, что касается перевариваемых углеводов, они представлены всеми классами. Полисахариды: крахмал (запасающий полисахарид растений), немного гликогена (запасающий углевод животных), когда едим мясо и печень. Олигосахариды: мальтоза в пиве и закваске вместе с изомальтозой, лактоза в молоке, сахароза (сейчас уже почти везде) и трегалоза (грибы). Моносахариды: редко встречаются в пище, но бывает.
Крахмал
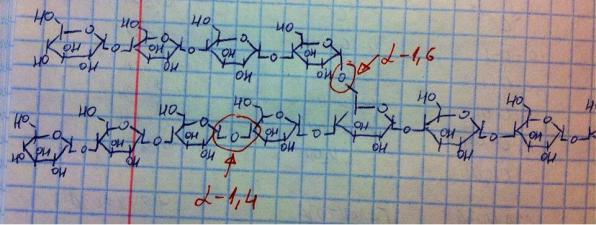
Основным углеводом пищи является крахмал, а остальные даже рядом не стояли, поэтому пищеварительная система настроена в основном на крахмал. Так что имеет ещё смысл обсудить строение крахмала – это запасающий углевод растений, полисахарид, разветвлённый гомополимер, мономер – глюкоза. Фрагмент крахмала представлен на рисунке выше, в крахмале идёт линейная цепь из глюкозы, связанной альфа-1,4 связью, но примерно на каждом 10-м мономере появляется ответвление, в котором две глюкозы соединяются альфа-1,6 связью. Итак, пищеварение. Уже в ротовой полости крахмал встречается с амилазой ротовой полости – эндогликозидаза, т.е. разрезает крахмал не с конца, а внутри и только по альфа-1,4 связи (банально потому, что их больше, чем альфа-1,6, поэтому фермент настроен на альфа-1,4). Работает фермент в слабощелочной среде, однако работает недолго, ибо еда во рту долго не задерживается и отправляется в желудок, в желудке нет ферментов для углеводов, зато внутри пищевого комка как бы всё ещё остаётся амилаза ротовой полости и
135
слабощелочная среда, поэтому переваривание идёт, пока в пищевой комок не проникнет желудочный сок, кислотность которого не позволит работать амилазе. Ротовая амилаза нарезает часть крахмала на декстрины – короткие полисахариды – а часть крахмала остаётся непереваренной, поэтому в рот поступал крахмал, а в 12-ти перстную кишку поступит крахмал и декстрины. В 12-ти перстную кишку выделяется из поджелудочной железы панкреатическая амилаза, которая действует точно так же, как и амилаза ротовой полости, только она активнее и времени у неё побольше, поэтому крахмал и декстрины она нарезает на мальтозу (минимальный кусок, который способны откусывать гликозидазы), изомальтозу (образуется в местах ветвления, ибо альфа-1,6 связь этим ферментом не гидролизуется) и мальтозоподобные олигосахариды 3-8 мономеров. Итак, переходим на уровень олигосахаридов, помимо того, что вышло из крахмала, в пищу попадают и другие сахариды – лактоза, сахароза, трегалоза. Олигосахариды расщепляются в основном ферментативными комплексами на наружной мембране энтероцитов – пристеночное пищеварение, плюс у этих ферментов есть дублёры в кишечном соке – полостное пищеарение, как и во рту, желудке, 12-ти перстной кишке. Первый комплекс – сахаразо-изомальтазный (окончания аза у фермента, оза у углевода – не путайте) состоит аж из 5-ти доменов: цитоплазматический, трансмембранный, внеклеточный (ножка) и непосредственно сахараза с изомальтазой, понятно, что они гидролизуют по альфа-1,2 связи сахарозу на глюкозу и фруктозу, а по альфа-1,6 связи гидролизуют изомальтозу на две глюкозы, однако данный ферментативный комплекс способен гидролизовать по альфа-1,4 связи мальтозу на две глюкозы, но делает он это с неохотой. Бета-гликозидазный комплекс гидролизует бета-1,4 связь в лактозе, делая из неё галактозу и глюкозу. Трегалазный комплекс по альфа-1,1 связи гидролизует трегалозу на две глюкозы. Глико-амилазный комплекс является экзогликозидазой, гидролизующей альфа-1,4 связи и превращая мальтозу и олигосахариды 3-8
в глюкозу. Итак, все углеводы расщепили до моносахаридов – можно всасывать. Фруктоза всасывается путём облегчённой диффузии, галактоза всасывается вторично-активным транспортом симпортом с натрием. Глюкоза всасывается обоими способами в зависимости от её концентрации в кишечнике: если её много в кишечнике, то по градиенту концентрации она поступит в энтероциты путём облегчённой диффузии, а если глюкозы в кишечнике мало, то она пойдёт против градиента концентрации в энтероциты вторично-активным транспортом. Из энтероцитов в кровь все углеводы попадают облегчённой диффузией.
136
Когда мы рассматриваем метаболизм углеводов и липидов, мы выделяем
•абсорбтивный и
•постабсорбтивный периоды.
Вабсорбтивный период происходит переваривание и всасывание углеводов (ну не только углеводов, просто в рамках этой темы нам нужно рассмотреть только углеводы), причём основной углевод пищи – крахмал, а он распадается на глюкозу, т.е. в абсорбтивный период мы получаем в основном глюкозу, да и остальные моносахариды способны превращаться в глюкозу, поэтому глюкоза – основной моносахарид нашего организма – вокруг неё всё крутится. Она является узловым метаболитом углеводного обмена. В итоге, в абсорбтивный период в кровь поступает большое количество глюкозы, больше, чем успевают съедать ткани, а концентрация глюкозы в крови – жёсткая константа – нельзя давать ей сильно вырастать, поэтому с помощью инсулина включаются пути утилизации глюкозы, один из основных – синтез гликогена в печени и мышцах, т.е. эти органы забирают из крови избыток глюкозы, однако глюкоза осмотически активная, если её будет слишком много в клетке, то клетка наполнится водой и лопнет, поэтому поступившая в клетку глюкоза тут же фосфорилируется и из неё синтезируют гликоген (осмотически не активный – его можно хранить в клетке).
Авот в постабсорбтивный период в кишечнике нет ничего, но ткани съедают глюкозу – её концентрация падает, в этом случае печень начинает мобилизацию гликогена, т.е. разрушает его до глюкозы и выпускает глюкозу
вкровь. Т.е. получается так, что ткани постоянно потребляют глюкозу крови, а кишечник вбрасывает в кровь глюкозу только периодически, зато устраивает тот ещё передоз – на лицо несогласованные вброс и расход глюкозы – эту проблему решает печень: когда глюкозы становится слишком много в абсорбтивный период, печень её запасает, снижая концентрацию до допустимой, а, когда глюкозы в крови не хватает, печень отдаёт свои запасы. Таким образом, печень отвечает за гомеостаз глюкозы, а первым и основным её механизмом является синтез и распад гликогена (запасающий углевод животных, структура, как у крахмала, но ветвление более обильное), конечно, это не единственные эффекторы в гомеостазе глюкозы – есть и ещё, но пока о синтезе и распаде гликогена. Обратите внимание, синтезируется и распадается гликоген и в мышцах, и в печени, но печень способна отдавать глюкозу в кровь, а мышцы – нет, поэтому-то печень в центре метаболизма и гомеостаза глюкозы.
Синтез гликогена
137
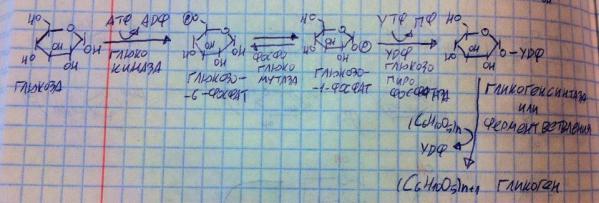
Итак, синтез гликогена.
Глюкоза попадает в клетку и тут же фосфорилируется с помощью глюкокиназы в 6-м положении. Глюкозо-6-фосфат осмотически неактивен, в отличии от глюкозы, поэтому клетке не грозит разрыв, плюс из-за данной реакции концентрация глюкозы в клетке практически равна нулю, а потому всегда меньше, чем в крови, что позволяет глюкозе входить в клетки по градиенту концентрации. Дальше на глюкозо-6-фосфат действует фосфоглюкомутаза (мутаза значит, что структура глюкозы не меняется, а переносится лишь с места на место группировка), она переносит фосфат в 1-е положение. Полученный глюкозо-1-фосфат взаимодействует с ферментом УДФ-глюкозопирофосфатой, который отщипляет ортофосфат от глюкозы и от УТФ, сшивает их в пирофосфат и удаляет, а оставшиеся глюкозу и УДФ связывает в УДФ-глюкозу. Дальше гликогенсинтаза убирает УДФ и насаживает глюкозу на гликоген с помощью альфа-1,4 связи либо это делает фермент ветвления, образующий альфа-1,6 связи. Мы специально переносили фосфат на 1-е положение, потому что именно с помощью гликозидного гидроксида этого атома углерода глюкоза образует связи. Заметьте, что гликогенсинтаза и фермент ветвления не работают без затравки, т.е. эти ферменты способны только наращивать гликоген, а не собирать его с нуля, по этой причине гликоген в нашем организме никогда не разрушается до конца. Вот так мы и получили гликоген. Ключевой фермент здесь – гликогенсинтаза, её активность управляется гормонами через химическую модификацию. Инсулин запускает дефосфорилирование гликогенсинтазы, делая её активной, а адреналин и глюкагон запускают её фосфорилирование и делают её неактивной. Процесс происходит в абсорбтивный период.
Распад гликогена
138
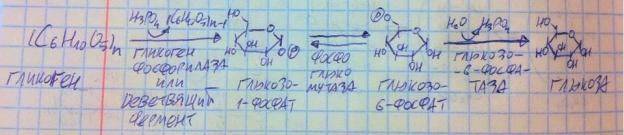
Распад гликогена происходит в постабсорбтивный период в печени (для нужд организма) и мышцах (обеспечение мышц энергией).
На гликоген действует гликогенфосфорилаза или деветвящий фермент,
в итоге альфа-1,4 или альфа-1,6, соответственно, разрываются, но не водой, а ортофосфатом, получается глюкозо-1-фосфат, который под действием глюкофосфомутазы превращается в глюкозо-6-фосфат, на этом мобилизация гликогена в мышцах кончается, продукт поступает в гликолиз, а в печени на него действует глюкозо-6-фосфатаза, которая гидролитически отщепляет ортофосфат, образуя чистую глюкозу, которая может выпуститься в кровь. Ключевой фермент – гликогенфосфорилаза, через инсулин дефосфорилируется, становясь неактивным, а через глюкагон и адреналин фосфорилируется, становясь активным. Т.е. заметьте, что распад и синтез гликогена управляются рецепроптно – когда включён один, другой выключен.
Синтез гликогена требует АТФ и УТФ на одну молекулу глюкозы, а распад гликогена идёт без затрат энергии.
Метаболизм глюкозы
Атеперь подробнее про метаболизм глюкозы и её гомеостаз. Глюкоза
–это жёсткая константа крови, она постоянно поддерживается на концентрации 3,3-5,5 ммоль/л. Почему так? Глюкоза в крови нужна – это факт, ведь это основной источник энергии в организме. Почему глюкозы в крови не должно быть слишком много? Глюкоза – осмотически активное вещество, если её в крови будет много, она вытянет воду из тканевой жидкости и притянет её в кровь, объём циркулирующей крови вырастет, вырастет и артериальное давление, помимо этого, при слишком высоких концентрациях глюкозы в крови, она начинает неферментативно присоединяться к белкам крови, изменяя их физико-химические свойства и нарушая тем самым их функцию. Почему глюкозы не должно быть слишком мало? Существуют системы, потребляющие глюкозу крови, а есть системы, обеспечивающие кровь глюкозой. И бывает такое, что при нагрузке ткани резко начинают больше потреблять глюкозу, уменьшая её количество в крови, это замечают системы, обеспечивающие кровь глюкозой, и начинают секретировать глюкозу в кровь, соответственно её потреблению тканями, однако эти системы отвечают с задержкой и если концентрация глюкозы в
139
крови изначально была бы крайне мала, то из-за этой задержки она могла бы на пару секунд падать до нуля, что как бы недопустимо. Второй аргумент
– разница концентраций между глюкозой в клетке и в крови – она должна быть достаточно велика, чтобы кровь входила в клетки с нормальной скоростью, ведь движущей силой транспорта посредством облегчённой диффузии является градиент (разница) концентраций. Вот поэтому глюкозу надо поддерживать в строго определённых концентрациях.
Итак, в абсорбтивный период много глюкозы поступает в кровь из кишечника, вся эта кровь устремляется по воротной вене в печень. Во всех тканях есть переносчики глюкозы ГЛЮТ разных модификаций, у печени они настроены на высокую концентрацию глюкозы, т.е. печень начинает потреблять глюкозу только, когда её уровень превышает предельно допустимые концентрации – заметная алиментарная гипергликемия – т.е. печень не отнимает глюкозу у других тканей, а забирает лишь излишки. Что же с излишками глюкозы делает печень? В первую очередь печень синтезирует из глюкозы гликоген, попутно работает гликолиз, ПДК, ЦТК, а там и ЦПЭ. Но если глюкозы слишком много, то вы всю в гликоген не засунете, да и не успеете, поэтому глюкоза отправляется в пентозофосфатный путь, где образуется НАДФН2, из ацетила-КоА (глицолиз + ПДК) и НАДФН2 строится холестерол и жирные кислоты, в гликолизе образуется дигидроксиацетонфосфат, из которого можно сделать 3-фосфо-глицерол, а из него и жирных кислот синтезируются триацилглицериды ТАГ (нейтральные жиры), которые отправляются по крови в ЛПОНП (липопротеины очень низкой плотности) в ткани, в том числе и в жировую. То есть, лишние углеводы превращаются в жиры. Всё, печень сделала, что могла, глюкозы стало значительно меньше в крови после печени, но, допустим, глюкозы всё ещё много, ибо мы те ещё сладкоежки, тогда включаются следующие линии обороны – жировая ткань и скелетная мускулатура – две эти ткани обладают по большей части ГЛЮТ-4 (переносчик глюкозы, который включается только инсулином, другие ГЛЮТы работают постоянно без инсулина). Эти ткани поглощают оставшиеся избытки глюкозы из крови, при этом в мышечной ткани синтезируется гликоген, а в жировой ткани синтезируются нейтральные жиры по тому же принципу, что и в печени. Все процессы утилизации глюкозы включает инсулин. Вот так в нашем организме утилизируется глюкоза путём синтеза гликогена и нейтральных жиров, первая линия обороны – печень (недаром воротную вену придумали от всего кишечника), вторая линия обороны – жировая ткань и скелетная мускулатура.
Теперь разберём постабсорбтивный период – глюкоза из кишечника не поступает, зато расходуется тканями, теперь, вместо кишечника, в кровь отдаёт глюкозу печень путём распада гликогена до глюкозы. Однако если
140