

Морфология растений / Нухимовский Е.Л. Основы биоморфологии семенных растений. III. Теория интегральной соматической эволюции
.pdf80 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
го, будучи причастным к биологии, — значит не дорожить чистотой и авторитетом науки, значит пренебрегать интересами и нуждами народа, которому призвана служить наука.
Мы глубоко убеждены, что работа Е.Л. Нухимовского при всей своей вычурной мудрёности, маскирующей идейное и научное убожество, представляет собой редкий образчик схоластики, метафизики и формализма в их современном, изощрённом обличии. Потенциальная опасность таких трудов гораздо больше, чем мы иногда склонны считать по своей снисходительности. Ибо лженаука остро заразна, а вспышки лженаучной заразы могут нанести, как свидетельствует история, колоссальный вред науке и обществу. Вот почему, чем скорее будет дана принципиальная оценка сочинению Е.Л. Нухимовского, тем меньшие материальные и моральные издержки понесёт наше общество.
Что же касается вопроса о соискании учёной степени, то, по нашему мнению, если бы инструкцией ВАК была бы предусмотрена возможность лишения на основании представленной работы уже имеющейся у соискателя степени кандидата наук, обсуждаемая диссертация с избытком удовлетворила бы необходимым для этого требованиям».
Что это за люди, которые, таким образом, обозначили своё присутствие в науке: гении, таланты, герои нашего времени… Нет же, это всего лишь типичные побрякушечники, их истинные лица; это их стиль, замешанный на жёлтой зависти — фирменный почерк пускать «пыль в глаза», «нести пургу»… Одна беда, их немало… и чем больше их будет, тем больше будут падать самолёты, тонуть корабли, взрываться электростанции и т.п. Мне тогда было 43 года, и эти люди отлично понимали «опасность», если его не остановить сейчас, он ещё многое сможет сделать в науке, а мы как были в ней, мягко говоря, «аморфной массой», так и останемся ею навсегда.
Профессор А.П. Пошкурлат тогда ответила побрякушечникам. «Получено 30 положительных отзывов, прослушали три положительных отзыва официальных оппонентов. Неужели все эти люди настолько не разбираются, что они дали положительные отзывы на такую безобразную работу и вот такие резкие выступления товарищей, они говорят против се-
бя…».
Да, я не стал профессором в отличие от многих моих коллег (для меня это невеликая потеря), и всю научную жизнь работал в должности младшего научного сотрудника (других для меня не нашлось, и это для меня не так уж важно), но я сделал в науке то, что доступно в ней лишь единицам, и впервые в мировой научной практике опубликовал трёхтомник по основам современной биоморфологии семенных растений (1997, 2002, 2020), объёмом около 2000 страниц, где, между прочим, рассмотрены особенности жизненных циклов 730 видов семенных фитобиоморф; эти книги вошли в фонды крупных библиотек мира и широко известны в России, а, кроме того, изданы десятки моих статей, среди которых пять опубликованы в одном из главных биологических журналов нашей страны
— в «Успехах современной биологии» (1980, 1984, 1986, 1987, 1988).
Чтобы не быть голословным о качестве своей диссертационной работы, я приведу её в полном объёме и без малейших изменений (см. стр. 626):
«Диссертация в виде научного доклада на соискание учёной степени доктора биологических наук» (2003, голосование 10:11). К сожалению, систе-
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
81 |
ма аттестации высших научных кадров далека от совершенства (я могу это всегда доказать) как во многих Учёных Советах, так и в ВАК’е, поэтому, если кому-то «сверху» угодно, то всегда будет обеспечено соответствую-
щее решение. Есть над чем поработать учёным следующих поколений. У меня нет сомнений, что решения о присуждении учёных степеней на защитах диссертаций, должны приниматься Учёными Советами только на основе отзывов оппонентов и ведущих учреждений, которые весьма авторитетно предварительно анализируют работы соискателей. Дискуссионные обсуждения по материалам диссертций всеми сторонами должны восприниматься только как информация к размышлениям и не более того, что реально будет перекрывать возможности всякого рода «побрякушечникам» влиять на результаты голосования Учёных Советов. Суть поднятых здесь проблем я понял давно и не стал слишком уж сокрушаться над тем, что происходило с моими докторскими диссертациями, тратить себя на борьбу «с ветряными мельницами» и попусту терять время на противоборство с побрякушечниками, поскольку оказалось, что проще всего их обойти (!), чтобы никогда, пока природой заведён в тебе рабочий механизм, не останавливаться на пути достижения более высоких целей в науке, а они далеки от конфронтаций с разного рода «побрякушечниками», «доброхотами», «пиарщиками» и т.п.
Продолжать дальше рассуждения на поднятую здесь тему бесполезно…, но я уверен в главном: мною прожита счастливая жизнь в науке, если оценивать её по истинным результатам.
2.5.3.Метод конвертируемого меротомирования, или, образно, — метод «грибной охоты»
Многие годы, экспериментируя и теоретизируя в биоморфологической работе с организацией семенных растений, я всё более и более стал воспринимать разнообразные методические приёмы в ней как искусство, в котором ты невольно, желая того или нет, начинаешь летать, образно говоря, «в облаках», отвлекаясь на какие-то моменты от объекта непосредст-
венного исследования, чтобы понять нечто большее, поэтому настойчиво начинаешь смотреть «по сторонам», менять углы обзора и высоты абстракции. В такой «игре» умодвижений можешь сталкиваться с чем-то но-
вым, чего раньше не замечал не только ты, но и другие, кто уже, на всё это, вроде бы, смотрел и не раз, и не два, а десятки, а то и сотни лет до тебя. И только сейчас в движениях именно твоих мыслей возникает картина, которую ранее не видел никто; в твоих методических блужданиях, наконец, свершается новаторское откровение, которое, возможно, само по себе не столь уж броско (нередко кто-то постарается его замять или совсем не заметить), но в нём есть сила, и в какое-то мгновение понимаешь, что это
только ты один обнаружил некоторые принципиальные «жемчужные» зёрна, способные прорастать и разрастаться в схемы, принципы и т.п., которые когда-то могут быть признаны открытиями.
Большинство фитобиоморфологов на вопрос, какие у семенных растений органы могут быть обозначены как основные, ответят единодушно: побеги и корни. Спорить на сей счёт бесполезно. Именно так считал и я, но только до некоторого, относительно недавнего момента, пока окончательно не убедился, что созданный мною методический приём, названный
82 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
по-научному методом конвертируемого меротомирования, или, образно,
— методом «грибной охоты», показал, что, если применить его к анализу организации семенных фитобиоморф, то, на самом деле, оажется не всё так уж и однозначно.
Для начала отвлечёмся несколько от биоморфологии и совершим путешествие в осенний лес с корзинкой в руках для грибной охоты, озадачив себя стремлением собрать побольше хороших грибов. Если только смотреть под ноги, то обычно на грибной охоте от этого будет мало пользы; здесь важно другое: умение максимально обозревать конкретный лесной участок, поэтому надо не только менять положение головы, но и туловища, наклоняясь, приседая и выпрямляясь, чтобы обозревать участок во все стороны и, что не менее важно, под разными углами. При таком подходе каждый отдельный лесной участок предстаёт перед нашими глазами по-разному, поэтому грибы находим там, где раньше прошли бы мимо.
Метод конвертируемого меротомирования нацелен наблюдать объект познания одного рода, например, основные органы семенных фитобиоморф, в единых природных явлениях (телах и процессах) под разными углами обзора и (или) с разной высоты абстракции, — по-разному, т.е. с
учётом иных организационных сторон — в разных форматах, что, таким образом, позволяет фиксировать естественные перемены (конверсии) в развитии природных явлений (семенных фитобиоморф, в частности) и, в соответствии с ними, производить в определённом исследовательском ключе членение (меротомирование) их единого признакового пространства на части. Используя этот метод, мы в главе 7 подробно разбираемся с разнообразием основных органов семенных фитобиоморф, определив их понятийные границы. Рассматривая основные органы в едином теле растительных организмов, как бывалый грибник, я стремился находить разные ракурсы при наблюдении за объектом исследования в своём стремлении так расчленить тело растительного организма, чтобы одни органы по найденным критериям проходили как основные, а другие — нет. При определении основных органов оказалось важным разграничивать их по статусным категориям: было обнаружено в организации семенных фитобиоморф не два, а десять типов основных органов, которые составили несколько групп из четырёх статусных категорий (подробнее см. главу 7).
2.5.4.Терминотворчество как методический атрибут развития наук, биоморфологии растений в особенности
Вряд ли кто-либо будет отрицать необходимость терминологиче-
ских исследований, и, пожалуй, все согласятся с Карлом Линнеем, которому принадлежит следующая фраза: «Без знания настоящего имени нельзя учинить ни коего описания, ни иметь понятия о вещах» («Словарь ботанический…», 1795). Важно знать не только правильное название видов, но и тех органов, различных структур и функций, посредством которых развиваются организмы (особи), представляющих содержание видов. Насколько важно точно определить название органов растений показывают некоторые парадоксы, отражённые в названиях видов, в первую очередь, систематиками из далёких прошлых времён, которые неправильно, на современном уровне знаний, распознали некоторые органы и, более того,
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
83 |
увековечили свои морфологические ошибки при описании новых видов. Во многом это не вина отдельных систематиков, а отражение уровня развития морфологии растений во времена, когда они жили и впервые определяли и называли новые виды. Например, в Израиле на побережье Средиземного моря по песчаным берегам произрастает интересный вид Crepis bulbosa (L.) Tausch; сейчас этот вид называют Aetheorhiza bulbosa (L.) Cass. (Danin, 1998, с. 211) (рис. 26). Этот вид образует подземные, иногда
ещё и приземные, побеговые клубни (клубнепобеги), которые в наше время никак не могут быть названы луковицами (bulbus). Не образуют клубней (tuberosa) некоторые другие виды с парадоксальными названиями, на-
пример, Thrincia tuberosa (L.) DC. (рис. 27), Lactuca tuberosa Jacq (рис. 28).
В реальности не бывают семенные растения «бесстебельными», поэтому остаётся только удивляться, когда некоторые систематики так называли отдельные виды (рис. 29).
|
10 |
|
у |
01к |
1кс |
|
||
|
пп |
|
|
|
43 39
61 |
78 |
74
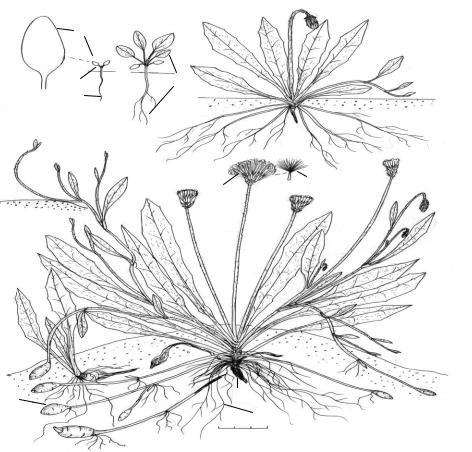
Рис. 26. Биографический портрет Ae heorhiza bulbosa (L.) Cass. (Crepis bulbosa (L.) Tausch.) [сем. Asteraceae (Израиль, недалеко от посёлка Зи-
хрон-Яаков, песчаный берег в 30–50 м от Средиземного моря, 22 III)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, закруглённые, 4–7 мм
дл., 2–3,5 мм шир., клиновидно суженные в черешок 2–5 мм дл.) розеточные столонообразующие клубнепобеговые компактнокорневищные жёлто-астроцветковые синорганизменные травянистые многолетники.

84 |
|
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
|
|
Б |
|
|
А |
у |
10 |
1кс |
|
||
|
В |
|
1к |
39 |
Д |
|
пп |
|||
|
|
||
|
|
43 |
5,15,18,21 |
91 |
71
31
к 
ск
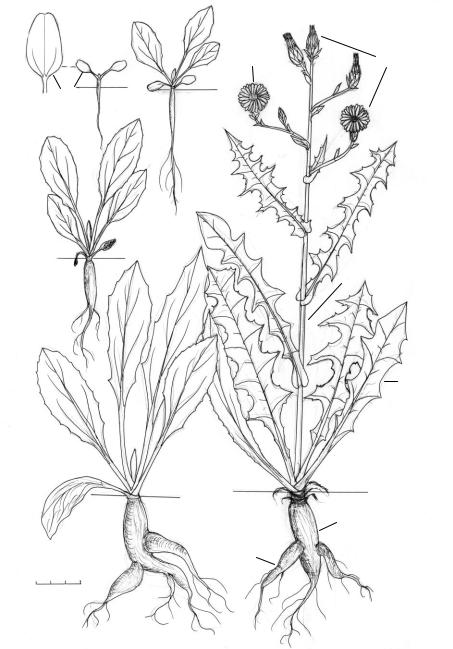
Рис. 27. Биографический портрет Thri cia uberosa (L.) DC. [сем. Asteraceae (в том же районе, что и рис. 26, горные склоны, 23 III)].
Биоморфотип: надземно-двусемядольные (семядоли продолговатые или продолговато-
эллиптические 5–8 мм дл., 1–2,5 мм шир., постепенно переходящие в короткие черешки) розеточные компактнокорневищные (главный корень обычно отмирает на 3–5 год жизни) гемисимподиальные жёлто-астроцветковые травянистые многолетники. Синонтогенезы продолжаются до 50–60 лет.
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
85 |
||
А |
Б |
Д |
|
|
|
||
|
43 |
91 |
|
у
10
В
 32 5,15,19,21
32 5,15,19,21
31
пп
72
73
Рис. 28. Биографический портрет Lac uca uberosa Jacq. [сем. Asteraceae
(Сосновые редколесья, луговины на горных склонах северного и центрального Израиля, заповедник «Neot Kedumim», 27 IV)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, часто выемчатые, 8–12 мм дл., 4–7 мм шир., клиновидно суженные в черешки 5–10 мм дл.) полурозеточные конодиальные мясисто-стержнекорневые светлофиолетово-астроцветковые унирепродуктивные обоеполые двулетние или малолетние (иногда живут 3–4 года) травы.

86 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
у
1л
1к
АБ
10 |
5,15,19,21 |
01к
1кс
Д
41
В
38
31
пп
ик
Г
55а
 11
11
4к 13
3л
72
Рис. 29. Биографический портрет Alcea acaulis (Cav.) Alef. [сем. Malvaceae
(Израиль, окрестности посёлка Зихрон-Яаков, разнотравье на горных склонах, 27 III)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, закруглённые, слегка
вдавленные с боков, сердцевидные в основании, 8–15 мм дл., 6–10 мм шир., на черешках 6– 15 мм) полурозеточные очерёднолистные преиндетерминантно-репродуцирующие (приот-
крытые, в средней и верхней частях побегов развиваются пазушные одиночные розоволепестковые цветки открытых фрондозных эфемерно-моноподиальных соцветий — кистей) однодомные каудексово-стержнекорневые травянистые многолетники.
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
87 |
2
10
у
1 |
7 |
|
сб
9 |
8 |
|
|
||
3 |
у |
|
9 |
||
4 |
|
|
11 |
5 |
|
13 |
12 |
|
пп |
||
|
||
6 |
|
|
14 |
|
|
15 |
8 |
11 сб
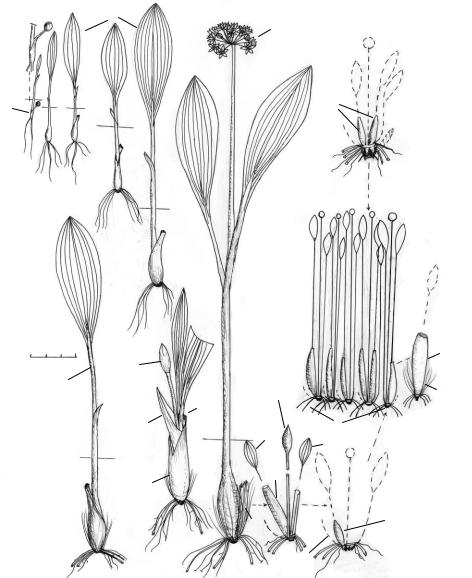
Рис. 30*. Биографический портрет Allium ursi um L. [сем. Liliaceae (Москва, Воробьёвы горы, парковая зона)].
1 — всход (IV), 2 — ювенильные растения (V), 3 — имматурное растение (20.V), 4 — молодое матурное растение (12.IV), 5 — многолетняя «семья» из нескольких потомковых растений от одной особи-предка семенного происхождения, или клон (25.VI), 6 — утол-
щённое основание зелёного листа, из которого образуется позднее фрондорезид, 7 — почколуковицы (15.VII) в составе единой солуковицы. 8 — почколуковица после отмирания зелёной пластинки листа (15.VII), 9 — бутон соцветия, 10 — соцветие (20.V), 11 — чешуя, 12 — эфемерный зелёный лист, 13 — «эфемероидный» (с отмирающей летом зелёной пла-
стинкой и не отмирающим до весны следующего года основанием) лист, 14 — фрондорезид в начале роста его зрелой почколуковицы рано весной следующего года, 15 — отмерший фрондорезид (24 V). Одно деление масштаба — 1 см, у — увеличено по сравнению с
масштабом. Морфология зимующей и весенней почколуковицы черемши до начала её отрастания — это корезид, или резидный («остаточный») побег, состоящий из фрондорезида (персистентного основания-остатка срединного листа), гемморезида (зимующей почки, которую прикрывает собой фрондорезид) и резида (персистентного стеблевого остатка от донца).
88 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
Работа с терминологией — обязательная часть всякого научного исследования, особенно биоморфологов, поскольку, в первую очередь, именно морфологи живых существ сталкиваются с необходимостью точно обозначить существование того или иного органа, находя для этого признаки, отличающие его от других органов. В методическом терминотворчестве уместно наилучшим образом сразу решить, а что такое означает само слово «термин». Термин, по моему мнению, — «это концептуальная динамическая система, характеризующаяся единством слова-этикетки (слова-символа) и понятия (объяснения этого слова в смысле, приемлемом для отражения явления-вещи, процесса определённого рода)» (Нухимов-
ский, 1997, с. 5). Такое понимание слова «термин» используется во всех моих работах. Насколько важна терминология для развития науки особенно ярко проявилось в медицине, где путём направленного терминотворчества удалось создать стройную систему знаний о множестве разнообразных органов и их частей в человеческом теле — анатомию человека. В исследованиях фитобиоморфолога обычно нет необходимости столь детально, как в медицине, описывать каждый элемент растительного тела, но всё-таки, чем больше мы знаем о деталях в существовании тех или иных
структур в организации растительных биоморф, тем более плодотворными могут стать наши практические и теоретические исследования. В своей биоморфологической деятельности мне удалось разработать и предложить немало терминов (резид, синорганизм, синонтогенез, персистентность, специкула, корнепобег и др.), некоторые из которых уже используют и другие морфологи (подробнее см. т. 1, 2).
В настоящей заметке есть смысл обратить внимание на то, как возникают новые термины в моей работе. Ответ простой: новые термины в науке обязаны возникать только по необходимости, когда без них становится неудобно работать дальше. Именно такое правило в терминотворчестве я принимаю, отлично понимая, что успешными смогут стать только действительно полезные, благозвучные и грамотно построенные термины. Приведу несколько примеров своего методического терминотворчества. Пожалуй, начну с черемши (Allium ursinum L.), поскольку биология этого
вида представляется мне интригующе интересной. Популяция этого вида, где проводились мои наблюдения, произрастает в лесопарковой зоне на Воробьёвых горах (Москва), здесь я её обнаружил случайно недалеко от места своего проживания. Возможно, она была занесена сюда человеком ещё в 50-е годы прошлого столетия, когда обустраивалась территория,
прилегающая к МГУ. Сейчас известная мне популяция черемши окрепла настолько, что успешно расширяет свою территорию, активно размножаясь и семенами, и вегетативно. Ботаники уже изучали черемшу и неоднократно описывали её биологию (Irmisch, 1850а; Серебряков, 1952; Van den Bergen, 1958; Старостенкова, 1978, и др.), поэтому я буду обращать внимание только на некоторые моменты из биоморфогении Allium ursinum.
Эфемероидные биоморфогенезы представители этого вида проходят по типу синонтогенезов (рис. 30), т.е. биоморфы, нередко сложные, — это
синорганизмы: сначала они развиваются как мероаклонисты (особи семенного происхождения чаще всего за 3–5 лет проходят стадии виргинильного этапа от проростка до начала отрастания первого цветоносного
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
89 |
побега, когда визуально фиксируется переход в матурный этап развития), а с образованием двух и более особей вегетативного происхождения (клонистов) становятся клонами. Побеговая система виргинильных особей нарастает моноподиально, точнее эумоноподиально (Нухимовский, 1986, 1997, с. 121), когда простая ось нарастает более одного года без отмирания (утраты) её верхушки».
В вегетационном сезоне у виргинильных (ювенильных и имматурных) особей, помимо стеблевого сближенноузлового, очень компактного донца и верхушечной почки, развиваются один нижний чешуевидный лист и один зелёный лист, основание которого несколько утолщается и сохраняет жизнеспособность до конца весны следующего года, а сама зелёная пластинка такого листа существует только до середины текущего лета, затем быстро желтеет и отмирает. Чешуя, утолщённое основание зелёного листа, донце и верхушечная почка таких растений составляют особое структурное образование (пробазовый орган, т.е. предшествующий базовому органу) — незрелую «почколуковицу». Термином «почколуковицы» удобно называть луковицеподобные почки, до начала образования из них соответствующих побегов видимого роста. Почколуковицы, например, развиваются у чеснока (Allium sativum L.), которые в текущем вегетаци-
онном сезоне формируются как незрелые почколуковицы, а после завершения сезона вегетации они превращаются в зрелые почколуковицы, зимующие вместе на одном донце в структуре единого базового органа — зрелой солуковицы. Термин «солуковица» (cobulbus) был предложен авто-
ром для обозначения базовых органов, напоминающих внешне луковицу, но, в отличие от неё, развивающих под покровом рано отмирающих, но долго сохраняющихся оснований зелёных листьев (или чешуй), пучок почколуковиц, которые по мере созревания этой системы теряют связь друг с другом (Нухимовский, 1997, с. 314). Я предлагаю применять термины «почколуковица» и «солуковица» также в отношении черемши.
В матурном этапе нарастание побеговой системы становится гемисимподиальным (с неежегодным, дифазным, т.е. во втором вегетационном сезоне, перевершиниванием), или моноподиально-симподиальным (под-
робнее см. т. 1, с. 121).
Персистентные (не отмирающие всё лето и осень, а затем зимующие до весны следующего года) основания акронекрозных, т.е. частично отмирающих сверху, зелёных листьев, которые образуются определённым образом у биоморф черемши, на мой взгляд, удобно назвать специальным термином. Здесь можно придумать разные слова, но я далеко ходить не буду и предложу новый научный термин «фрондорезид» [от лат. frons, frondis — листва + residuum — остаток (Забинкова, Кирпичников, 1957; Кирпичников, Забинкова, 1977)]. Это, в нашем случае, базальный, утол-
щённый, остаток листа, сохраняющий жизнеспособность более одного вегетационного сезона (персистентный), после утраты им какой-либо верх-
ней части (у черемши зелёная пластинка листа отмирает в середине лета). В более широком значении фрондорезид — это любой зелёный лист или только базальный остаток его, сохраняющий жизнеспособность до весны следующего года или дольше. Фрондорезиды, наряду с гемморезидами и резидами, развиваясь совместно, образуют в единстве резидные побеги,