

Морфология растений / Нухимовский Е.Л. Основы биоморфологии семенных растений. III. Теория интегральной соматической эволюции
.pdf240 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
в отдельные стороны бинариев. Разнообразию бинарных союзов в организации семенных фитобиоморф уделим соответствующее внимание.
Мысли о том, что в организации живой природы (шире во всей Природе) действует принцип бинарного эволюционизма, навели меня на рассуждения о взаимоотношениях личного (своего) и неличного (не-своего) в
организации семенных фитобиоморф и в их эволюции (т. 2, гл. 2.5). Давайте именно с этого материала и начнём наш путь фактического рассмотрения принципа бинарного эволюционизма. Этот материал особенно важен, на мой взгляд, поскольку на его основе построена концепция компромиссов в эволюции растительных биоморф, которая прошла красной нитью через всё содержание второго тома нашей монографии.
5. 2. 1. Синдром вегеторепродуктивной бинарности
Из предыдущего материала мы установили, что бинарность — это проявление в целостности какого-либо объекта-ресурса (тела или нетела)
двойственности (двоичности), каждая из сторон (солитариев) которой взаимодополняя друг друга в развитии, тем не менее, несёт в себе ещё и противоречия, разрешение которых сопровождается разнообразными событиями внутри и (или) вне носителей двойственности (бинариев) где-то без компромиссов, а где-то с компромиссами между соответствующими
сторонами. Получается, на самом деле, что с позиции бинарного принципа любые ресурсы (тела и нетела) обладают какими-либо формами бинарно-
сти, которые надо и захотеть, и суметь распознать, увидеть. Бинарный принцип эволюции — это рассмотрение объекта эволюции (тела или нетела) через призму присутствия в нём некоторых форм бинарности, в которых фиксируются противоречивые события из отношений отдельных сторон (солитариев) определённых бинарных систем (бинариев), и, в итоге, устанавливается результат их взаимодействий в диапазоне развития соответствующего бинария и его отражения в эволюции целого объекта. В каждом объекте-ресурсе может содержаться много двойчатых систем — би-
нариев, стороны которых (солитарии) находятся в организационном единстве, но и в противоречиях, нередко глубоких. Бинарии, или бинары, будучи всегда системами из двух атрибутивных солитариев, обнаруживаются в объекте под определёнными углами обзора и высот абстракции. Поскольку целостность объекта эволюции многоформативна в бинарном отношении, то и рассматривать такие объекты можно и нужно по-разному в
каждом отдельном бинарном формате. Меросоматическая организация и меросоматическая эволюция — это части некоторых целых явлений, а значит это могут быть только солитарии в развитии внутри соответствующих бинариев, поэтому можно рассматривать особенности некоторой солитарной эволюции в организации каких-либо объектов. Когда идёт
речь об эволюции целого объекта в формате действия определённой би-
нарности, то это уже уровень интегральной (ценосоматической) эволюции.
Бинарии проявляют себя через состояния, сменяющиеся или не сменяющиеся, объектов эволюции в отдельные отрезки времени их существования (бытия) или через одно состояние, которое сопровождает бытие
Глава 5. Бинарный принцип в соматической эволюции |
241 |
объекта с момента возникновения до конца его бытия, т.е. весь цикл его существования. Эволюция, являясь непрерывным процессом, сопровождается множеством бинарных событий как в онтогенетическом, так и филогенетическом развитии.
Стремление осуществить размножение является проблемой, целенаправляющей онтогенетическое эволюционное развитие биоморф. Стремление разрешить противоречия между вегетативом и репродуктивом является проблемой, целенаправляющей филогенетическое эволюционное развитие. Бинарностей много, но мы здесь обратим внимание, дополняя предыдущие исследования (т. 1, 2), на ту из них, которая в эволюции семенных фитобиоморф стала одним из атрибутов (нормативных обязанностей) и решающим фактором в определении направленности эволюции этих объектов в разнообразных условиях окружающей среды, что позволило мне говорить об этом явлении как о синдроме (от греч. syndrome — сте-
чение) вегеторепродуктивной (вегетативно-репродуктивной) бинарно-
сти. Обычно комбинацию симптомов, характеризующих определённую болезнь, определяют в медицине как синдром чего-то. В последнее время
этот термин постепенно расширяет свой смысл, поскольку многое в живой природе, что не является болезнью, имеет некоторую склонность вести себя подобно болезни, проявляясь комбинацией симптомов, характеризующих какое-либо явление, в нашем случае, например, это один из видов
бинарности. Здесь, пожалуй, будет уместно заметить, что биоморфы, когда присутствуют в развитии в одном признаковом «поле» (пространстве), то являются носителями определенных бинарностей, например, абио-
биосоматической бинарности, которая по соответствующим приоритетным признакам рассматриваемого явления определяет их, эти тела (биоморфы) как абио-биосоматические бинарии. В другом признаковом «по-
ле» биоморфы являются носителями иных бинарностей, например. вегеторепродуктивной, которая позволяет определять эти биоморфы под соответствующим углом обзора и определённой высоты абстракции уже как вегеторепродуктивные бинарии. Далее можно продолжать, если это необходимо, различать в признаковых пространствах биоморф другие бинарности, например, гено-феносоматическую бинарность и т.д.
На мой взгляд, проявление синдрома вегеторепродуктивной бинарности в организации биоморф вообще, семенных фитобиоморф в том числе, — это величайшее достижение витаэволюции (эволюции жизни) на всех уровнях её (гипобиоморфном, биоморфном и метабиоморфном, т.е. в мире эврибиоморф, или форм жизни в широком смысле).
Возникновение вегетатива в морфогенезе биоморф — это сигнальный признак того, что где-то несколько позже организация биоморф уст-
ремится к развёртыванию другой своей бинарной стороны — способности размножаться, причём семенные фитобиоморфы более всего в своём развитии целенаправленны на репродуктивное размножение. Цель — это ещё не само счастье, а только дорога к нему, что афористически метко подчеркивает неизбежный факт: путь к реализации репродуктивной функции тернист и требует от любых биоморф больших вегетативных усилий.
Во-первых, оптимально удобно расположить вегетативное тело со-
ответствующей биоморфы, устремлённое к достижению репродуктивного

242 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
размножения, чтобы, тем самым, будучи в окружении других живых существ (и друзей, и врагов), наиболее весомо поучаствовать в сохранении не-своей (неличной) жизни — в сохранении вида, что является приори-
тетом в жизни каждой биоморфы.
|
|
у |
Б |
Д |
|
А |
43 |
||
|
|
|||
|
|
|
|
|
01к |
|
|
10 |
|
|
|
|
|
2кс
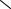 33а
33а
пп
Рис. 116. Биографический портрет Sisymbrium erysimoides Desf. [сем.
Brassicaceae (Израиль, западное побережье Мёртвого моря, разреженное разнотравье на горных склонах, пересохшие русла ручьёв, 9 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые, 4–6
мм дл., 2–3 мм шир., клиновидно суженные в короткие черешки 2–4 мм дл.) полурозеточные очерёднолистные семидетерминантно-репродуцирующие (полузакрытые, образуются верхушечные открытые брактеозно-абрактеозные кистевидные соцветия) жёлто-капустно-
цветковые однодомные стержнекорневые эвриэфемерные травы.
Во-вторых, биоморфы обязаны поначалу проявить максимум уси-
лий, чтобы создать вегетативное тело, которое будет способно в нужный момент и в нужном месте приступить к реализации репродуктивных усилий.
В-третьих, вегетатив биоморф должен быть оптимально пластичным, чтобы как-либо трансформируясь выдерживать неудобства в отно-
шениях с конкурирующими организмами и иным агрессивным давлением внешних факторов.
Так уж сложилось в морфогенезах (онтогенезах или синонтогенезах) каждой биоморфы, а это не что иное, как проявление текущей (морфогенезной) эволюции, обязательное соматическое развёртывание вегеторепродуктивного бинария происходит стадийно, и каждый из солитариев (отдельных сторон единого бинария) строго отрабатывает свою миссию в биоморфогенезе (жизненном цикле биоморфы).
Первый вегетативный солитарий, или вегетосолитарий (v-солита-

Глава 5. Бинарный принцип в соматической эволюции |
243 |
рий, если сокращённо) может в развитии биоморфы остаться единственным в своём образе, если, после развёртывания репродуктивного солитария, или re-солитария (от лат. re — снова + production — производить), а с ещё большим сокращением будет r-солитарий, развитие бинария пойдёт
далее по нисходящей и закончится элиминацией биоморфы. Когда в биоморфогенезе проявляется развитие только одного вегеторепродуктивного бинария и жизнь биоморфы продолжается не более одного года, то имеет место эвриэфемеризм (рис. 116–135). Если в биоморфогенезе стадии вегетативного и вегетативно-репродуктивного развития будут два и более раз
повторяться, сменяя друг друга, и жизнь биоморфы будет продолжаться более одного года (рис. 136–143), то проявляется бирепродуктивная или мультирепродуктивная персистентность (многолетность) развития.
А 10 |
Б |
43
01к
у
2,15,21
Д |
29 |
|
28 |
2кс
31,33а
71 
Рис. 117. Биографический портрет Diplotaxis acris (Forssk.) Boiss. [сем.
Brassicaceae (там же, что и рис. 116, разреженное разнотравье на горных склонах, 10 III)].
Биоморфотип: надземно-двусемядольные (семядоли сильно модифицируют от узкояйцевидных до яйцевидных, чаще продолговато-яйцевидные, закруглённые 5–8 мм дл., 2,5–4
мм шир, клиновидно суженные в черешки 3–5 мм) полурозеточные очерёднолистные се- мидетерминантно-репродуцирующие (полузакрытые верхушечными открытыми брактеоз- но-абрактеозными кистевидными соцветиями) фиолетово-капустноцветковые однодомные
стержнекорневые эвриэфемерные травы.

244 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
10
01к
40,70
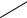 2,15,21
2,15,21
2кс
пп
Рис. 118. Биографический портрет Lappula spinocarpos (Forssk.) Ascherson
[сем. Boraginaceae (Израиль, окрестности посёлка Сде-Бокер, крутые склоны каньона в пустыне Негев, 30 III)].
Биоморфотип: надземно-двусемядольные (семядоли от эллиптических до яйцевидных,
закруглённые, 6–10 мм дл., 3,5–5 мм шир., опушённые, клиновидно суженные в черешки 2–5 мм дл.) безрозеточные очерёднолистные детерминантно-репродуцирующие (закрытые, на верхушках побегов развиваются фрондозно-фрондулёзные эфемерно-полисимпо- диальные соцветия) фиолетово-бурачниковоцветковые однодомные стержнекорневые тра-
вянистые эвриэфемеры.
10 |
Ж |
|
01к
1кс
2к
40
пп
Рис. 119. Биографический портрет Anchusa aegyptiaca (L.) DC. [сем.
Boraginaceae (там же, что и рис. 118, 25 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые,
опушённые, 8–12 мм дл., 4–6 мм шир., на черешках 2–3 мм дл.) полурозеточные очерёднолистные детерминантно-репродуцирующие (закрытые, на верхушках побегов развиваются фрондулёзно-брактеозные эфемерно-полисимподиальные завитковые соцветия) светложёлто-бурачниковоцветковые однодомные стержнекорневые травянистые эври-
эфемеры.

Глава 5. Бинарный принцип в соматической эволюции |
245 |
у
10
43
01к
1кс
34 |
2,15,21 |
пп
Рис. 120. Биографический портрет Anthemis leucanthemifolia Boiss. et Blanche [сем. Asteraceae (Израиль, окрестности Тель-Авива, овраги, обочины дорог, 14 IV)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые, 3,5–
6 мм дл., 2–3 мм шир., сидячие) безрозеточные или полурозеточные очерёднолистные се- мидетерминантно-репродуцирующие (полузакрытые, на верхушках главного побега и его
ветвей развиваются белолепестковые корзинки) однодомные стержнекорневые сорные травянистые эвриэфемеры.
А |
10 |
у Б |
1,14,18 |
В |
сб |
|
|||||
|
|
|
|
Д
2,15,21
 2,19,21
2,19,21
 35
35
пп
Рис. 121. Биографический портрет Anthemis pseudocotula Boiss. [сем.
Asteraceae (там же, что и рис. 118, 30 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые, сидячие. 4–6 мм дл., 3–4 мм шир.) полурозеточные очерёднолистные семидетерминантно-
репродуцирующие (полузакрытые, верхушки побегов заканчиваются белолепестковыми корзинками) однодомные стержнекорневые травянистые эвриэфемеры.

246 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
10 |
42 |
01к |
2,15,20,21 |
|
у 
Рис. 122. Биографический портрет Reseda boissieri Mueller [сем. Resedaceae
(там же, что и рис. 116, 12 III)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные или продолговато-
яйцевидные, закруглённые, 6–8 мм дл., 2–3 мм шир., на черешках 2–5 мм) безрозеточные очерёднолистные семидетерминантно-репродуцирующие (полузакрытые, с верхушечными открытыми фрондулёзно-брактеозными соцветиями, составляющими вместе метёлку из кистей) бело-резедоцветковые однодомные стержнекорневые травянистые эвриэфемеры.
10 |
|
|
1кс |
01к |
|
у |
2,15,19,21 |
|
43 |
пп
Рис. 123. Биографический портрет Reseda decursiva Forssk. [сем. Resedaceae (там же, что и рис. 118, 30 III)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, закруглённые, 6–8 мм дл., 3–4 мм шир., 2–5 мм черешки) полурозеточные очерёднолистные семидетерминантно-
репродуцирующие (полузакрытые, верхушечные соцветия — открытые брактеозные кисти) однодомные стержнекорневые травянистые эвриэфемеры.

Глава 5. Бинарный принцип в соматической эволюции |
247 |
10
у 
01к
1кс
2,15,20,21
пп
 71
71
Рис. 124. Биографический портрет Knautia integrifolia (Coulter) Greuter (K. bidens Sm. Lindley). [сем. Dipsacaceae (Израиль, окрестности посёлка Зи- хрон-Яаков, редколесье и поляны на горных склонах, 11 IV)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые, 8–14
мм дл., 4–6 мм шир., клиновидно суженные в черешки 3–5 мм) безрозеточные супротивнолистные семидетерминантно-репродуцирующие (полузакрытые, на верхушках разветвлён-
ного главного побега развиваются открытые брактеозные компактные соцветия — головки) однодомные стержнекорневые травянистые эвриэфемеры.

248 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
10 |
1кс |
|
43 |
|
39 |
|
у |
|
2к |
2к |
|
Рис. 125. Биогрфический портрет Rumex cyprius Murb. [сем. Polygonaceae
(там же, где и рис. 118, 27 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические или продолговато- эллиптические, или продолговато-яйцевидные, закруглённые, 7–10 мм дл., 2,5–4 мм шир.,
клиновидно суженные в черешки 2–4 мм) полурозеточные очерёднолистные семидетерми- нантно-репродуцирующие (полузакрытые, на верхушках побегов развиваются открытые
брактеозные кисти, что создаёт эффект неполной закрытости биоморф сверху их репродуктивной сферой) розово-гречиховоцветковые (округлые листочки околоцветника при пло-
дах розовые) стержнекорневые травянистые эвриэфемеры.
|
А |
|
Б |
|
|
|
|
||
01к |
10 |
|
43 |
|
|
|
|||
у |
1кс |
пп |
||
|
||||
Д |
|
|
|
71
Рис. 126. Биографический портрет Matthiola aspera Boiss. [сем. Brassicaceae
(там же, что и рис. 118, 27 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые 4–7
мм дл, 3–4 мм шир., на черешках 2–4 мм) полурозеточные очерёднолистные семидетерми- нантно-репродуцирующие (полузакрытые, на верхушках побегов развиваются открытые брактеозные кисти розово-фиолетоволепестковых цветков) однодомные стержнекорневые
эвриэфемерные травы.

Глава 5. Бинарный принцип в соматической эволюции |
249 |
||
|
А |
Б |
Д |
у |
10 |
1кс |
42 |
|
|
|
|
В
2,19,21
1к |
31,34 |
|
Рис. 127. Биографический портрет Ragadiolus stellatus (L.) Gaertner [сем. Asteraceae (там же, где и рис. 124, 1 IV)].
Биоморфотип: надземно-двусемядольные (семядоли нитевидные, мясистые, слегка за-
острённые, 15–30 мм дл., 0,5–1 мм шир., сидячие) полурозеточные очерёднолистные семи- детерминантно-репродуцирующие (полузакрытые, на верхушке главного побега развивается метельчатое соцветие из корзинок) эфемерно-полисимподиальное (по типу нарастания осей соцветия) жёлто-астроцветковые однодомные эвриэфемерные травы.
|
10 |
31 |
|
|
|
|
|
|
|
40,70 |
Др |
|
|
|
|
у |
|
10 |
|
|
|
1кс |
|
|
|
40 |
|
|
01к |
|
|
пп 5,19,21
Рис. 128. Биографический портрет Cynoglossum creticum Miller [сем. Boraginaceae (Израиль, заповедник «Neot Kedumim» и окрестности города Ариэль, разнотравье на открытых горных склонах, соответственно 15 III и
25 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические или яйцевидные,
опушённые, закруглённые, 12–18 мм дл., 6–9 мм шир., клиновидно суженные в короткие черешки, почти сидячие) полурозеточные очерёднолистные детерминантно-репродуци-
рующие (закрытые, на верхушках главной оси и ветвей унирепродуктивного побега развиваются монохазиальные, эфемерно-полисимподиальные соцветия типа завиток) фиолетово-
бурачниковоцветковые однодомные стержнекорневые травянистые двулетники. Др — об-
щий вид взрослых растений (Feinbrun-Dothan, Danin, 1991, с. 534).