

Морфология растений / Нухимовский Е.Л. Основы биоморфологии семенных растений. III. Теория интегральной соматической эволюции
.pdf90 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
или корезиды (см. с. 89). Роль фрондорезидов в жизни биоморф черемши переоценить невозможно: она одна из главных на эфемероидном пути их образа жизни. Именно такие «эфемероидные обрубки» только некоторых, по своему особенных, зелёных листьев — немного утолщённые остаточные основания их, сохраняющие жизнеспособность до весны следующего года (фрондорезиды), позволяют считать базовый (персистентный, т.е. живущий не менее одного года) орган не «ложной луковицей» (Филимонова, 1959), а иначе, что, естественно, требует объяснения. Фрондорезид у черемши выполняет роль депо для накопления воды и питательных веществ, поэтому именно он является одним из основных структурных элементов почколуковицы (gemmobulbus), которая в текущем сезоне своего
образования является «незрелой», а в период относительного зимнего покоя — «зрелой» (терминологические новации на сей счёт см. подробнее т. 1, с. 314). Я не уверен, что термин «почколуковица» новый, поскольку смысл его лежит как бы на поверхности. Почколуковица у особей черемши живёт около одного года, а затем сменяется другой почколуковицей, причём у виргинильных особей при моноподиальном нарастании, а у матурных — при гемисимподиальном нарастании. Персистентные листовые элементы у черемши до цветения растений включают только 1 фрондорезид, а у взрослых особей образуется или один, или два, очень редко (!) (при значительном углублении в почву) больше (3) фрондорезидов. Почколуковица неполовозрелых особей цельная. Некроз постепенно охватывает донце солуковицы и далее перемещается на самые нижние части каждой почколуковицы; к середине весны, или несколько позже, некроз разрушает и почколуковичные перезимовавшие фрондорезиды, которые уже выполнили свою функцию, обеспечив из перезимовавшей почки отдельных почколуковиц начало весеннего роста цветоноса и новых боковых побегов (одного, реже более). Матурные (половозрелые) особи у черемши бывают, по определению М.М. Старостенковой (1978), «двухосными» (с одним боковым побегом), когда развёртываются 2 зелёных листа, или «трёхосными», если зелёных листьев — 3 (с двумя, очень редко более, боковыми побегами). Базовый орган в последнем (трёхосном) варианте (рис. 30) особенный, что, на мой взгляд, важно зафиксировать терминологически, подчёркивая, как бы между прочим, что ничего «ложного» в организации тела живых существ не бывает. У трёхосных особей черемши развиваются дифрактные (от лат. difractus — разорванный, рваный), или раз-
рывные солуковицы. У таких солуковиц, в результате отрастания цветоноса, а в дальнейшем ещё и его некроза исчезает общее «покрывало» для потомковых («сестринских» по М.М. Старостенковой, 1978) почколуковиц, которые становятся свободными, будучи сначала частично связанными через дряхлеющее донце, а к концу весны или к началу лета они полностью отрываются друг от друга. Дифрактность (разрыв, в результате роста, одной временно целой солуковицы) и образование двух, редко более, свободных потомковых почколуковиц (незрелых в текущем вегетационном сезоне и зрелых — зимой) — важный показатель жизнесохранительной способности черемши, как вида, успешно поддерживать свои популяции не только семенным, но и вегетативным размножением. Полный распад в начале лета дряхлеющей структуры родительского донца дифрактной со-
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
91 |
луковицы приводит у особи с двумя почколуковицами к образованию клона. Образование у простых биоморф (организмов) черемши особых органов, отличных по многим признакам от луковиц, когда на одном донце формируется только одна почколуковица, а реже две, иногда даже три парциальных почколуковицы (дифрактная, или разрывная солуковица) заметно расширяет разнообразие базовых (персистентных) органов семенных фитобиоморф. По типу базовых органов Allium ursinum L. проявляет двойственность — это аклонально-почколуковичные и клонально-
почколуковичные растения, а в целом, приоритетным для этого вида является биоморфотип почколуковично-клонирующих травомноголетних растений (рис. 30). Allium ursinum — это один из примеров весьма ориги-
нального явления в соматической эволюции организации семенных фитобиоморф — викароперсистентности (Нухимовский, 1997, с. 316).
Немало сторон организации фитобиоморф, которые ещё не нашли в ботанике для себя надёжного терминологического определения. К числу таковых, например, относится термин «корнеплод», который активно применяют как в обиходной практике, так и в научной литературе, хотя все отлично понимают, что здесь нужен другой термин, поскольку никакого «плода» на самом деле не образуется, а возникает корне-стеблевая доб-
рокачественная опухоль нижней части тела некоторых розеточных и полурозеточных однолетних, двулетних и реже иных малолетних растений. Здесь назрела необходимость просто-напросто найти более удачное слово-
этикетку как обязательной части термина, пожалуй, мудрствовать не приходится. На мой взгляд, хорошо известное понятие найдёт достойное выражение и отражение в качестве научного термина со словом-этикеткой (ярлыком) «келоид» [keloidum (от греч. kele — опухоль + eidos — подобный, имеющий вид)]. Келоид, будучи доброкачественным разрастанием базальных стебле-корневых частей, образуется как у некоторых однолет-
них или двулетних, чаще всего розеточных и полурозеточных стержне-
корневых, растений [Raphanus sativus L., Beta vulgaris subsp. esculenta
Guerke, Daucus sativus (Hoffm.) Roehl. и др. (рис. 31)].
Иногда у многолетников образуются корневые клубни, помогая таким фитобиоморфам решать проблемы запасания воды и минерально-
органических веществ, например, Filipendula vulgaris Moench, Phlomoides tuberosa (L.) Moench (т. 1, с.316), Erodium crassifolium L’Her. (рис. 32).
Особый интерес вызывают доброкачественные разрастания некоторых многолетников, контрактильная корневая активность которых создаёт специализированный подземный клубневидный орган с гипокотильно-
корневой структурой; предлагаю называть его «ризокелоидом» (рис. 33, 34). Этот запасающий орган хорошо адаптирован в структуру эфемероидов, а выше этого органа обычно развивается базовый (многолетний побеговый) орган, в частности каудекс (рис. 33, 34).
Я морфолог, если вижу структуры, функции, явления, которые не имеют своего названия, и их, чтобы лучше понять, о чём идёт речь, приходится предварительно с разных сторон многословно описывать, то это своеобразный сигнал, чтобы задуматься о вероятностной полезности именно здесь применить терминотворческие действия. Так, например, мною были предложены новые термины: резид, синорганизм, синонтоге-
92 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
нез, конодий, персистентность, специкула, корнепобег, эусимподиальность, гемисимподиальность, викароперсистентность и др.
Терминотворчество, на самом деле, — это ответственный методический приём, особая процедура научной работы, к которой исследователь может и должен прибегать по необходимости, когда появляется навязчивая потребность сказать ясно и кратко о многом, используя для этого всего лишь одно, или реже, два «заветных» слова-этикетки (обязательные части
всякого термина), за которым обязано стоять глубоко осмысленное понятие. Я вовсе не призываю кого-либо устраиваться в кресла поудобнее и
пытаться навешивать «ярлычки» на разнообразные явления и понятия, полагая, что и таким образом можно поучаствовать в развитии науки. На мой взгляд, важно глубоко осознавать необходимость соответствующих терминотворческих новаций, а ситуации нередко складываются весьма непростыми, даже больше, они бывают коварно-интригующими и нередко
требуют усилий многих поколений учёных (для этого достаточно присмотреться, например, к тому, как изменялась ботаническая терминология со времён Карла Линнея).
Когда-то, ещё в студенческие годы, я увлёкся разработкой проблемы
организации каудексовых растений. В эти годы я активно участвовал в экспедициях (Московская область, Калмыкия, Казахстан), много месяцев провёл в библиотеках и особенно в Музее книги Российской государственной библиотеки, что, в конце концов, позволило конкретизировать термин «каудекс», а заодно, на основе эмпирического материала и абстрактного мышления, удалось разработать модель разнопризнаковой и мультилинейной организации биоморф семенных растений (Нухимовский, 1968, 1969а, б, 1970, 1971, 1997). В этих исследованиях родился новый для науки термин «резид» (от лат. residuum — остаток, reses, idis — остающийся) — стеблевая часть побега, которая стала скелетной (двух–, трёх–,
многолетней) после завершения им видимого роста и (или) репродуктивного развития. Побеги, каркас которых составляют резиды — резидные, или персистентные (подробнее см. т. 1, гл. 6.2). У термина «резид» теперь своя судьба, поскольку он вызвал интерес и его уже используют морфологи в своих исследованиях. Есть идеи применять термин «резид» шире его исходного смысла (Мастинская, 1978; Жмылёв и др., 2005; Костина, 2009, и др.). Иногда предлагают применять этот термин только в отношении отдельных групп семенных фитобиоморф, например, «резид — многолетнее одревесневшее основание надземного побега у многолетних стержнекорневых и кистекорневых трав» (Коровкин, 2007). Я по-прежнему считаю,
что термин «резид» — это персистентный, или зимующий стеблевой остаток от побега, завершившего свой активный рост после окончания вегетационного сезона, зимующего и (или) далее сохраняющего жизнеспособность более одного года у любых персистентных (дву–, многолетних) семенных фитобиоморф. Если резид рассматривается вместе ещё и с другими не отмирающими зимой частями побега — зимующими почками (гемморезидами) и зимующими (когда они есть) листьями или отдельными частями — зимующими остатками листьев (фрондорезидами), то это резидный побег, или корезид (от лат. co — вместе + резид).

Глава 2. Методология и методика на пути к эволюционному теоретизированию |
93 |
|||
|
А |
Б |
|
|
|
31 |
|
|
|
|
|
|
|
|
у |
10 |
43 |
Д |
|
|
|
|
||
Г2,15,19,21
1кс пп
В
32 
Рис. 31 А. Биографический портрет Be a vulgaris L. в условиях естественного произрастания [сем. Chenopodiaceae, Израиль, заповедник «Neot Kedumim», открытые горные склоны, 1 V)].
Биоморфотип: надземно-двусемядольные (семядоли продолговатые или чаще продолго- вато-яйцевидные, 15–20 мм дл., 5–7 мм шир., постепенно сбегающие в короткие черешки)
полурозеточные очерёднолистные стержнекорневые однолетние травы («келоид», или «корнеплод» не образуется).

94 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
 а
а
а
 а
а
б б
б |
кд |
кд |
|
пп
кд |
в |
в |
|
в |
|||
|
|
3
1 2
Рис. 31 Б*. Схема
строения запасающих органов у некоторых цветковых растений (по Рау-Rauh, цит. по
Ал. А. Фёдорову, М.Э. Кирпичникову и З.Т. Артю-
шенко, 1962, с. 49).
1 — Свёкла сахарная
(Beta vulgaris L. var.
altissima Rössig.; 2 —
Свёкла кормовая (Beta
vulgaris L. var. rapacea Krass.); 3 — Свёкла
обыкновенная (Be a vulgaris L.). а — ли-
стья, б — гипокотиль, в — корень, пп — уровень почвы; кд — келоид (расположение органа, для которого в этой работе предложен новый термин).
С момента появления определённых терминов в науке у них идёт отсчёт уже их бытия, их истории и, образно говоря, время в далёкой перспективе обычно всё расставляет по местам.
Терминотворчество иногда завлекает. Однако, если хорошо присмотреться, то создаётся впечатление, что где-то даже без него вполне
можно и обойтись. Можно, но нужно ли? Приведу пример из материалов этой книги, здесь и сейчас я предложу использовать новый термин, который вовсе не обязателен для широкого внедрения, но, тем не менее, на мой взгляд, он «не паразит», поскольку в некоторых случаях весьма уместен, даже очень необходим. Такой термин просто обязан удачно вписываться в материал и создавать эффект, способствующий наиболее полному осознанию происходящих событий с объектом исследований, в нашем случае — семенными фитобиоморфами. Я предлагаю термин «витараптация» (от лат. vita — жизнь, raptus — резкое движение, взрыв), которая бывает веге-
тативной, когда распускаются (образно «взрываются», естественно биотически) вегетативные почки, или репродуктивной, когда происходит цветение и плодоношение, и смешанной (вегетативно-репродуктивной), когда
оба эти события свершаются одновременно. В медицине, например, используется термин «взрыв тоскливости» — raptus melancholicus (Арнау-
дов, 1979). Можно ещё шире посмотреть на Мир, на нашу Вселенную вообще, чтобы обратить внимание на общеизвестные факты: многие события, которые происходят в безбрежных космических пространствах, сопровождаются физическими взрывами разного рода, например, вулканов, звёзд, галактик и т.д. Биотические взрывы («витараптусы») — результат развития соматической организации фитобиоморф, сопровождающиеся

Глава 2. Методология и методика на пути к эволюционному теоретизированию |
95 |
массированным выбросом, разрядом или витараптацией энергии вегетативного и (или) репродуктивного развития, достигшей критической массы: весеннее пробуждение почек и (или) цветение, плодоношение. Кто-то, возможно, скажет: стоит ли таким деталям, прямо-таки мелочам… уделять
внимание. Но Вы, уважаемый читатель, далее по ходу книги, присмотритесь к этому термину и сами примите решение был ли он уместен в этой работе и есть ли у него перспектива на будущее применение в ботанической науке. Главное, конечно же, время, которое спокойно всё расставляет по своим местам: чему более бывать или не бывать.
Б |
В |
|
А 10 |
2к |
|
1кс |
пп |
|
01к |
94 |
|
у |
||
|
||
Д |
|
|
дс |
91 |
|
5к |
|
|
2,5,15,19,21 |
||
29 |
|
|
64 |
|
|
|
70 |
|
31,35 |
|
|
72 |
|
|
73 |
|
|
64 |
|
|
71 |
|
|
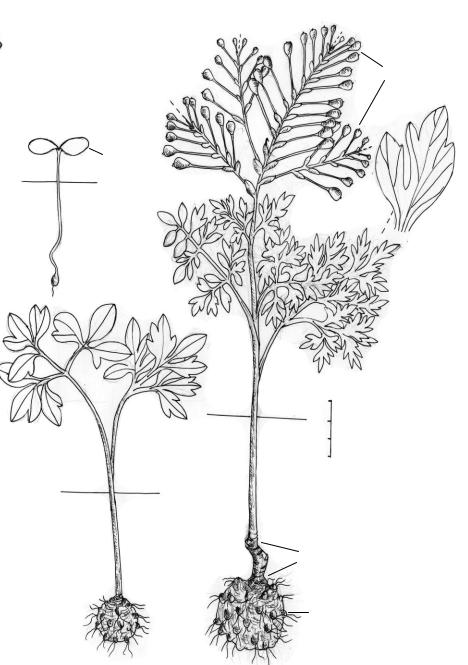
Рис. 32. Биографический портрет Erodium |
crassifolium L’Her. [сем. |
|
Geraniaceae (Израиль, каменистая пустыня Негев, окрестности посёлка Сде-Бокер, 25 III)].
Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, при основании серд-
цевидные, 6–8 мм дл., 4–6 мм шир., нередко неравнобокие, у других обе лопасти семядоли супротивные) полурозеточные эфемерно-полисимподиальные (в репродуцирующей зоне) гемисимподиальные (в вегетативной основе) клубнекорневые (ризокелоидные) фиолетово- геранецветковые стержнекорневые многолетние травы. Онтогенез продолжается 10–15, редко до 20 лет.
96 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
43
10
 64
64
пп
55а
64
Рис. 33. Биографический портрет Leontice leontopetalum L. [сем.
Berberidaceae (там же, что и рис. 32, 25 III)].
Пример каудексово-ризокелоидного эфемероидного травянистого многолетника.

Глава 2. Методология и методика на пути к эволюционному теоретизированию |
97 |
|
Б |
В |
|
31 |
|
|
А |
|
|
|
|
|
11 |
|
|
10 |
43 |
|
12 |
|
|
13 |
у |
|
Г
Д
пп |
55а |
 64
64
Рис. 34. Биографический портрет Scorzonera judaica Eig [сем.
Asteraceae (там же, что и рис. 32, 25 III)].
Пример эумоноподиального каудексово-ризокелоидного травянистого
многолетника. Высокая контрактильность в процессе развития подземных органов буквально затягивает в клубнекорневую зону весь гипокотиль.
98 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
2.6. Визуально-фитобиографический метод
Эволюционное теоретизирование в отношении мира живых существ обычно осуществляется при обязательном накоплении достаточного фактического материала, в котором, на мой взгляд, важное место должны занимать разнообразные сведения о жизненных циклах биоморф, т.е. организмов и синорганизмов, — об их биографике. Жизненный цикл биоморфы — биоморфогенез (онтогенез или синонтогенез), а обобщённые сведения о биоморфогенезах определённого таксона, например, вида, — биоморфогения, или биографика соответствующего таксона (онтогения или синонтогения). Раздел биоморфологии, изучающий биоморфогенезы, — биографика (биоморфогения). Биографика, как уже было подчёркнуто ранее (т. 2, гл. 4.2), способна как самостоятельно решать задачи индивидуального и кондивидуального развития растений, так и быть основополагающим разделом (сердцевиной) ещё более крупной науки — биоморфологии. В развитие биографики растений (фитобиографики) внесли свой вклад фитоморфологи разных поколений (Irmisch, 1850а, б, 1859, 1860,
1861, 1874; Warming, 1884, 1908, 1909, 1916; Velenovsky, 1905, 1907, 1910, 1913; Богдановская-Гиенэф,1926; Goebel, 1928, 1931; Troll, 1937, 1939, 1954, 1964, 1969; Пошкурлат, 1941, 1962, 1975, 2000; Серебряков. 1952,
1954, 1962, 1964; Игнатьева, 1958, 1964, 1965; Серебрякова, 1959, 1962, 1971; Уранов, 1960, 1973, 1974; Борисова, 1961; Голубев, 1962, 1965;
Csapody, 1968; Диагнозы, 1983; Смирнова, 1987; Татаренко, 1996; Биоло-
гическая флора Московской области, 2000; Коломейцева, 2004, 2005, 2007; Современные подходы к описанию структуры растений, 2008; Актуальные проблемы современной биоморфологии, 2012; Онтогенетический атлас растений, 2013, и многие другие). Впервые наиболее последовательные представления о жизненных циклах и дифференциации их на возрастные состояния сформировались в работах Т.А. Работнова (1945, 1946, 1947, 1949. 1950, 1960, 1964, 1974, и др.), что позволяет считать его одним из основоположников биографики растений.
Существуют разные способы передачи фитобиографической информации, что мы ранее отразили в трёх типах (таблично-рисуночном, пове- ствовательно-рисуночном и рисуночном), в которых решающее значение
отводится рисункам с детальными условными обозначениями к ним. Таким образом удаётся кратко и последовательно отражать различные стороны становления жизненных форм (организмов и синорганизмов) от их рождения и до смерти (т. 1). Уже сложился набор обязательных вопросов, на которые желательно отвечать при изучении биоморфогенезов (т. 1, гл. 7, 9; т. 2, гл. 4) и дан список обязательных вопросов программы-минимум для таких исследований (т. 2, с. 340–341).
Долгое время при изложении особенностей жизненных циклов семенных фитобиоморф использовались довольно просторные описания, где подробно пересказывались события, которые наглядно отражались в рисунках и (или) схемах, и чаще всего многое было понятно без дополнительных разъяснений такого рода. Например, подробно описывалось, что семена, развиваясь весной, набухают и главный корень, прорывая семенную оболочку, устремляется в глубь почвы, а гипокотиль, дуговидно изги-
Глава 2. Методология и методика на пути к эволюционному теоретизированию |
99 |
баясь, выносит семядоли над поверхностью почвы, где они вскоре развёртываются и постепенно увеличиваются в размерах, достигая определённых размеров… и т.д. Так биоморфологи работали в 20 веке, когда происходило становление биографики — науки о жизненных циклах (биоморфогенезах) биоморф (организмов и синорганизмов), но к настоящему времени обстоятельства изменяются; прошло уже немало десятилетий с моментов зарождения биографики, когда ещё было приемлемо писать, например, о том, что гипокотиль удлиняется после начала прорастания семени и, дуговидно изгибаясь, выносит на поверхность почвы семядоли, где они, несколько поднимаясь над ней, развёртываются, приобретая зелёную окраску, и многое подобное из подробностей.
В современной фитобиографике, на мой взгляд, нужно освобождаться в описаниях от всего тривиального, обычно понятного без разъяснений для всех специалистов. Полагаю, что в XXI столетии в фитобио-
морфологии решающим методом передачи информации об индивидуальном (простые биоморфы) и кондивидуальном (сложные биоморфы) развитии семенных растений станет рисуночный метод, который удобно называть ещё и «визуально-биографическим». Визуализация (от лат. visualis —
зрительный) — «представления явления или процесса в наглядной форме, удобной для зрительного восприятия» (Большой сл. иностр. слов, 2012, с. 149). Суть метода, в нашем случае визуально-фитобиографического мето-
да, в том, что основное содержание в накоплении и передаче биоморфогенезной информации закладывается в рисунки (может дополняться схемами и фотографиями), снабжаемые подписями к ним, включающие, в первую очередь, подробные условные обозначения. На основе прочтения информации, заложенной в рисунки, составляются резюме, где выделяются разнообразные биоморфотипы, характеризующие возрастные состояния и целостное развитие фитобиоморф в конкретных условиях их произрастания. В основном я уже давно, с разной степенью последовательности, работаю в биоморфологии растений с использованием визуально-фито-
биографического метода (т. 1, 2).
В основном здесь, чтобы не быть голословным, приведу краткие описания биоморфогении в форме биографических портретов большой группы видов семенных растений по визуально-фитобиографическому методу (рис. 35–63), которые мне удалось изучить в заповеднике «Галичья
гора» (Липецкая область), где я побывал на конференции по гидрофитам (август, 2005), весьма успешно организованной А.В. Славгородским и А.И. Кузьмичёвым при содействии руководства заповедника, после которой я задержался на несколько дней, чтобы посмотреть биоморфологию некоторых местных растений. Как за три дня удалось изучить биографику более 30 видов? Во-первых, был продолжительный летний рабочий день
— от зари до заката солнца при хорошей погоде. Во-вторых, было доста-
точно популяций с разновозрастным растительным материалом; трудности иногда возникали с поиском проростков и всходов, что было естественно, поскольку наблюдения велись в конце лета, но было достаточно семенного материала. По возвращении, я ещё некоторое время выращивал
в Московской области на дачном участке из семян молодые растения, что восполнило недостающие данные. В методическом отношении использо-