

2. Система імунітету людини та її вплив на перебіг вірусної інфекції
2.1. Імунна система та її реакція на вірусну інфекцію
Більшість патогенних вірусів проникає в організм хребетних тварин через слизові оболонки або в кровотік (укус тварин, комах та ін.). Отже, шкіра та слизові є першим бар'єром на шляху вірусної інфекції. До механізмів неспецифічної резистентності відносять біологічні бар'єри, клітинні і гуморальні фактори, фізіологічні функції систем і органів. Специфічна резистентність забезпечує взаємодію Т-і В-лімфоцитів, Т-лімфоцитів і макрофагів (табл. 6).
Таблиця 6.
Механізми захисту організму ссавців від ураження вірусами
Види захисту |
Неспецифічні |
Специфічні |
Зовнішні бар’єри (шкіра, слизові оболонки) |
Механічний бар'єр (непроникність шкіри і слизових оболонок). Секреторний бар'єр (підвищена кислотність вмісту шлунка). Рух війок. Відсутність рецепторів для вірусів на поверхні клітин, що виключає адсорбцію. |
Імуноглобуліни (IgA) |
Гуморальний захист |
Присутність неспецифічних противірусних інгібіторів (термолабільних β-інгібіторів) і термостабільних α - і ɣ-інгібіторів, що покривають поверхню віріона і блокуючих адсорбцію (наприклад, під час грипу). Фагоцитоз (гранулоцити (мікрофагоцити) і тканинні макрофаги). Система комплементу. Різке підвищення температури організму (лихорадка). |
Імуноглобуліни (IgD, IgE, IgM, IgG) |
Клітинний захист |
Інтерферон виходить з уражених вірусом клітин і захищає від інфікування сусідні, здорові клітини. Фагоцитуючі клітини: нейтральні гранулоцити, еозинофільні гранулоцити, мононуклеарні фагоцити, натуральні кілери. Відсутність в клітинах ферментів, необхідних для репродукції віруса. Синтез ендогенного інтерферону, що продукують уражені вірусом клітини |
Клітинний імунітет (Т-лімфоцити і макрофаги) |
Не пошкоджена, нормальна, шкіра і слизові оболонки активно виконують бар'єрну функцію. Слизові оболонки носа і верхніх дихальних шляхів непроникні для багатьох вірусів, за винятком вірусу грипу. Зміна стану організму (стрес, голодування, переохолодження і ін.). Знімає опірність слизових оболонок і збільшує їх проникність для мікроорганізмів. Важливу роль грає фіксація мікробів на поверхні слизової оболонки і подальше їх механічне видалення. Мікроби і пилові частинки при вдиху придавлюються до поверхні слизової оболонки, а потім видаляються за допомогою чхальних і кашльових рефлексів, руху миготливого епітелію, злущування епітеліальних клітин з поверхні слизової оболонки, перістальтічних скорочень бронхів. Змивання мікробів зі слизової оболонки дихальних шляхів викликається рясним відділенням слизу та інших секретів, що володіють бактерицидною дією. При попаданні в організм збудників через шлунково-кишковий тракт (кишкові інфекції) першим бар'єром на їх шляху стають слизові оболонки ротової порожнини і слина. Слина містить фактори бактерицидної і бактеріостатичної дії (лізоцим, бактеріолізін, IgA). Взаємодіючи з секреторним IgA, лізоцим викликає лізис резистентних до нього бактерій, але противірусної активністю не володіє. Крім лізоциму в слині, відокремлюємо носоглотки і в сироватці крові містяться інгібітори гемаглютинації, перешкоджаючі взаємодії вірусів з чутливими клітинами. Сильним бар'єром є шлунок.
Лімфатичні вузли - наступний бар'єр на шляху збудника інфекційної хвороби. У новонароджених лімфатичні вузли не диференційовані. Бактерії погано затримуються в синусах і слабо захвачуються макрофагами, що пояснює легкість розвитку в цьому віці генералізованих і септичних форм інфекціі. Генералізація інфекційного процесу особливо легко наступає в умовах недосконалої нейро-ендокринної регуляції і високої здатності до проникання гемато-енцефалічного бар'єру. У новонароджених речовина мозку недостатньо диференційована і дуже добре забезпечується кров'ю, тоді як відтік крові істотно важко через незавершеність розвитку венозної системи. Створюються сприятливі умови для акумулювання токсичних речовин, особливо бактеріальних і вірусних токсинів, чим і пояснюється частіше виникнення у дітей раннього віку токсичних форм інфекційних захворювань.
Серед неспецифічних чинників захисту найважливіше значення має фагоцитоз. Розрізняють дві популяції клітин циркулюючих, що фагоцитують, в крові гранулоцити (мікрофагоцити) і тканинні макрофаги.
Мікрофагами є нейтрофіли, а макрофагами - моноцити. Фагоцитарна функ-ція нейтрофілів починає формуватися вже на 15-25 тижні внутрішньоутробного розвитку.
Дитина народжується з досить вираженою фагоцитарною активністю лейкоцитів. Але це стосується лише процесу хемотаксису, руху і поглинання, тоді як переварювання поглинених мікробів ще значно понижене і досягає величин дорослого на 6-12 місяці життя. Система фагоцитозу може бути генетично дефектна, а крім того ряд мікроорганізмів, такі як пневмокок, клебсиелла, гемофільна паличка та ін., взагалі не піддаються завершеному фагоцитозу. У цих випадках перебіг хвороби може бути особливий важким з розвитком деструктивних процесів або сепсису.
Макрофаги можна розглядати як мікроеталон неспецифічної резистентності організму. Якщо вірус не розмножується в макрофагах, організм є резистентним.
Комплемент - система, що включає, принаймні, 18 сироваткових ків, які активирующихся в каскаді реакцій. Здійснює лізис сенсибилизирован¬ных антитілами клітинних антигенів, обусловли¬вает реакцію імунного прилипання, бере участь в опсонізації бактерій, вірусів, прискорюючи їх фаго¬цитоз.
Дитина народжується з низькою активністю сис¬теми комплементу (близько 50% від рівня активно¬сти дорослого). Проте, вже на першому місяці життя, тивність системи комплементу швидко зростає і досягає рівня дорослої людини до 6-ти місячному віку.
Інтерферон - низькомолекулярний білок з противірусними властивостями, виробляється клітинами організму, інфікованими вірусом. Інтерферон продукується параллельно з репродукцією вірусу. Найбільш активними ндукторами інтерферону є міксовіруси і ентеровіруси. Іноді його продукція може випереджати або, навпаки, відставати від розмноження вірусу. Усі інтерферони діють головним чином як противірусні чинники. До найбільш вивчених ефектів відносяться:
активація макрофагів;
активація натуральних кілерів;
пригноблення вірусної реплікації.
Противірусна дія інтерферону проявляється на внутрішньоклітинному етапі репродукції вірусу. Здатність до продукції інтерферона генетично детермінована і передається у спадок, вона істотно залежить від віку. Відразу після народження здатність до утворення інтерферону відносно низька, потім вона підвищується, досягаючи максимуму до 12-16 років.
Інтерферон виробляють практично усі клітини організму, причому процес виробництво інтерферону починається вже у вхідних воротях інфекції. Тому результат зустрічі мікро- і макроорганізму певною мірою буде залежити від активності интерфероногенеза вже на ранніх етапах інфекційного процесу.
Разом з противірусною дією, інтерферон пригнічує внутрішньоклітинне розмноження малярійних плазмодіїв, хламідій, рікетсій. Показано, що у присутності інтерферону клітини є стійкіші до дії екзо- і эндотоксинів.
Інтерферон посилює фагоцитоз, пригнічує трансформацію клітин онкогенними вірусами, гальмує зростання пухлинних клітин, підвищує цитотоксичность лімфоцитів. У дітей першого місяца життю інтерферон гальмує диференціювання нервової тканини.
Якщо вірус проникає через бар'єр неспецифічної резистентності, тоді починають діяти специфічні чинники захисту - гуморальні (імуноглобуліни) і клітинні (сенсибілізовані лімфоцити). Під час вірусних інфекцій вирішальна роль принадлежить клітинному імунітету.
1. Специфічні імунологічні реакції пов' язані в довгий ланцюг з Т- і В-системою імунітету. Ці клітини спільно з макрофагами здійснюють найголовніші типи імунологічного реагування, включаючи вироблення антитіл і накопичення сенсибілізованих лимфо¬цитов, що розпізнають і видалити чужерідні субстанції.
2. Нейтралізуючі вірус антитіла перешкоджають проникненню вірусу в клітину. Механізми взаємодії вірусних антигенів з антитілами не досить вивчені, проте розглядаються два можливі варіанти взаємодії антигенів з антитілами:
антитела взаємодіють з поверхневими білками вірусних оболонок, блокуючи їх здатність зв'язуватися з рецепторами клітинної поверхні;
антитіла змінюють просторову орієнтацію вірусних антигенів і при цьому
а) порушується взаємодія вірусу з клітинним рецептором;
б) порушується злиття вірусних оболонок з клітинною мембраною і проникнення вірусного нуклеопротеїду у цитоплазму.
Виунокомпетентності клони лімфоїдних клітин дозрувають в різний час. Раніше за все з'являється здатнвсит до імунної відповіді на антигени вірусів, жгутиковий антиген сальмонел, антиген стафілокока.
В процесі дозрівання імунної системи плоду має значення проникнення деякої кількості різних антигенів через плаценту і внутріутробна підготовка лімфоїдних клітин плоду до бактерійних і вірусних антигенів, що найчастіше всього зустрічаються.
Особливо швидке дозрівання імунної системи дитини відбувається після народження, у зв'язку з масовим мікробним обсіменінням і багатократним збільшенням антигенного навантаження. У цьому процесі велику роль грають умовно-патогенні бактерії, що колонізують верхні дихальні шляхи і шкірні покриви, особливо шлунково-кишковий тракт.
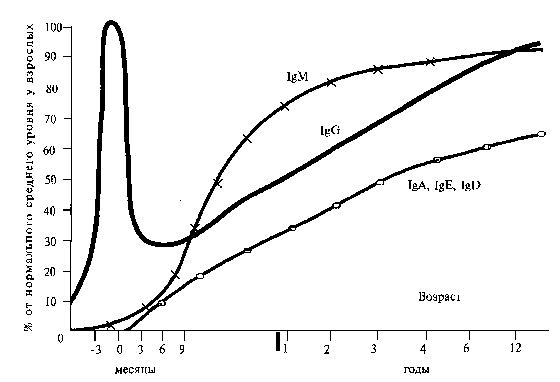
Про стан специфічного иммунологічного реагування прийнято судити по вмістув сироватці крові імуноглобулінів класів (A, E, D, G, M) (рис. 28)
При народженні
Рис. 28. Зміна рівнів змісту імуноглобулінів в сироватці крові, при народженні і в постнатальному періоді
Імуноглобуліни М складають близько 10% усього пулу імуноглобулінів в організмі, до його складу входять антитіла проти грамнегативних бактерій (шигелл, сальмонелл, та ін.), Частково проти виру¬сов, розчинних антигенів і токсинів. Антитіла класу 1gМ не проходять через плаценту, тому новонароджена дитина може захворіти шигеллезом, сальмонельозом, респіраторною вірусною інфекцією та ін.
Антитіла, що відносяться до 1gМ мають високу аглютинуючу активністю і виконують важливу роль в антимікробному імунітеті. Синтез антитіл 1gМ починається вже в перший тиждень життя дитини, швидко наростає, досягаючи рівня дорослого на 12 - 24 міс. життя.
Імуноглобулін G - складає до 80% усіх імуноглобулінів. У його складі розрізняють чотири підкласи: 1gG1, 1gG2, 1gG3 і 1gG4. Клас 1gG містить велику частину противірусних антитіл, а також антитіла до бактерійних токсинів, полісахаридів, Клітиної стінки, капсул грампозитивних бактерій, до типоспецифічного М-протеіну стрептокока. Підкласи 1gG1 і 1gG3 можуть індукувати фагоцитоз, а підкласи 1gG1, 1g G2 і 1g G3 здатні фіксувати комплемент. Найважливішою особливістю усіх під класів 1gG являється їх способ-ность проходити через плацентарний бар'єр. Процес переходу 1gG через плаценту особливо активно осуществля¬ется в останні тижні вагітності, тому зміст 1gG у недо-ношенных може бути низьким або повністю бути відсутнім. У тих же випадках, коли народжується доношена дитина зміст 1gG в його пуповинній крові відповідає рівню такого у матері або навіть перевищує цей рівень (см. рис. 22). Тому новонароджена дитина не захворює, як правило, кором, епідемічним паротитом і дифтерією
Відразу після народження починається процес катаболизму пасивно отриманих 1gG і рівень його поступово знижується, досягаючи мінімальних значень на 6-9 міс. життя Саме в цьому віці дитина найбільш сприйнятлива до багатьох вірусною і бактерійною інфекційною захворювань.
Синтез власних антитіл класу 1gG, хоча і виявляється ще внутріутробний, проте після народження цей процес блокується высо¬ким рівнем трансплацентарних антитіл цього класу. У міру катаболізму трансплацентарних антитіл наростає синтез власних імуноглобулінів класу 1gG, але рівня дорослого чоло¬століття вони досягають лише на 5-6 році життя дитини.
Імуноглобуліни А складають за об'ємом близько 15% усіх імуноглобулінів сыво-ротки крові. Характерна присутність імуноглобуліну А в жіночому молозиві, слині, сльозах, носовому і бронхіальному секреті, а також на слизовій оболонці про¬лочке кишковика. Ці иммуноглобулины грають вирішальну роль в місцевому імунітеті, перешкоджаючі інвазії бактерій і вирусів в організмі. Але, оскільки 1gА не проходять через плаценту, дитина народжується з майже повною їх відсутністю. Цим можна пояснити особливу сприйнятливість новонароджених і дітей перших місяців життя до гострих респіраторних вірусних захворювань.
Важливо враховувати, що недостатність секреторних імуноглобулінів у новонароджених і дітей перших місяців життя компенсується грудним вигодовуванням. Показано, що молозиво і свіже жіноче молоко мцстять велику кількість 1gА.
Оцінюючи динаміку наростання основних класів імуноглобулінів в процесі розвитку і зростання дитини, можна бачити, що найбільша імунологічна незахищеність відзначається у віці від 6 місяців до 1,5 - 2 років. Саме у цьому віці дитина повністю втрачає пасивний материнський імунітет, тоді як здатність до вироблення власних антитіл знаходиться лише у стадії розвитку, далеко не досягаючи активности дорослої людини. Цим пояснюється легка сприйнятливість дітей цього віку до ГРВІ (гострих респіраторних вірусних інфекцій), шлунково-кишкових захворювань, іншим вірусним і бактерійним инфекціям.
Процес дозрівання імунної системи може бути порушений у зв'язку з внутріутробним інфікуванням дитини. У цих дітей синтез імуноглобулінів починається до народження. При цьому особливо значущо підвищується синтез имуноглобулинів М. Отже, високий зміст 1gM у новонародженого можна вважати найважливішим показником наявності природженої інфекції.
Після зіткнення з антигеном В-лімфоцити певного клона починають проду-кувати специфічні антитіла (імуноглобуліни). Функціональна активність В-лімфоцитів регулюється спеціальними Т-клітинами і гуморальними чинниками, такими, як ІЛ- 1 (інтерлейкін). При продукції антитіл раніше усього відбувається секреція IgM; В-лімфоцить потім діференциюються у плазматичні клітини, утворюючи клітини, що синтезують інші класи імуноглобулінів (IgG, IgA, IgЕ, IgD). Деякі популяції В-лімфоцитів не диференціюються в плазматичні клітини і стають В-клітинами пам'яті, які обумовлюють розвиток негайної реакції антитіла з повторно виділеним антигеном.
Система противірусного імунітету має деякі особливості, в порівнянні з імунітетом протибактерійним або протипухлинним:
1. В першу чергу спрацьовують інтерферони і Т-килерні клітини .
2. Віруси виявляють слабку антигенну дію на В-лімфоцити і для їх активування, проліферації (розмноження) і диференціювання потрібна участь Т-хелперів.
3. Противірусну дію мають α і β-інгібитори.
α-інгибітор входить до складу α-глобулинів, перешкоджає адсорбції вірусів на клітинній мембрані. -інгібітор входить до складу -глобулінів, пригнічує розмноження орто- і параміксовірусів.
4. Т-цитотоксичні лімфоцити і інші килерніе клітини знищують будь-які клітини, інфіковані чужими антигенами.
5. Противірусні антитіла, адсорбуючись на вірусних рецепторах, унеможливлюють прикріплення вірусу на мембрані клітини.
6. Проти внутрішньоклітинних форм - вірусів -- антитіла не ефективні.
Схема взаімодії клетин у противірусному імунітеті подана на рис. 26.
Р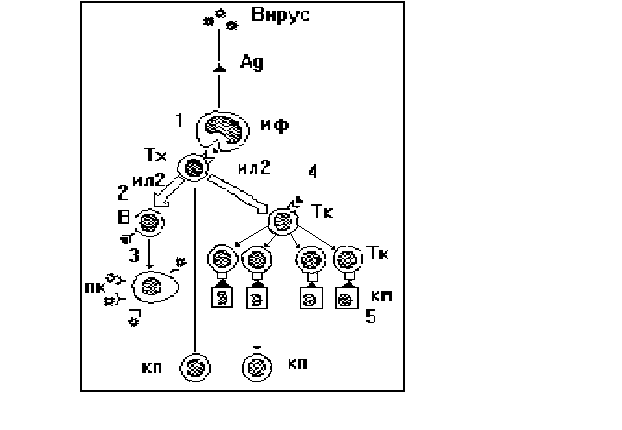 ис.
29.
Взаємодія
клітин у специфічному імунітеті
ис.
29.
Взаємодія
клітин у специфічному імунітеті
1 - представлення і розпізнавання вірусного антигена (Ag) за участю макрофагів (мф) і Т-хелперів (Тх);
2 - розпізнавання Ag(антигену) Т-лімфоцитами, їх активація продуктами Тх (изолейкінами - ІЛ- 2);
3 - формування клона продукуючих антитіла плазматичних клітин (пк) і продукція віруснейтралізуючих антитіл;
4 - формування клона Т-кілерів;
5 - розпізнавання Тк антигенів вірусу на мембрані клітин -"мишеней" (км);
6 - формування клітин пам'яті (кп).
Імунітет можна розділити на природжений (видовий) і придбаний.
Придбаний імунітет у свою чергу можна підрозділити на природний (як наслідок перенесеного захворювання), так і штучний. Штучний імунітет може бути активним (після вакцинації) і пасивним. Пасивний імунітет у свою чергу подрозділяється на природний (у разі передачі антитіл від матері дитині через плаценту) і штучний (як наслідок введення в організм імунних сироваток). Пасивний природний імунітет зберігається від 2 до 6 місяців, а пасивний штучний - до 2 тижнів.
Найбільш надійним і перевіреним засобом зниження захворюваності гострими інфекційними хворобами є активна імунізація (профілактичні щеплення). В результаті активної імунізації створюється антитоксинний, антиінвазивний або нейтралізуючий ефект в імунному органіизмі. Деякі імунізуючі агенти дають повний захист упродовж усього життя, інші - часткову а треті повинні застосовуватися повторно, через певні інтервали часу.
Планове проведення активної імунопрофілактики призвело до різкого зниження захворюванності багатьма інфекційними хворобами і зробило можливою ліквідацію такої особливо небезпечної інфекції, як натуральна віспа. Це досягнення стало результатом здійснення програми Всесвітньої організації охорони здоров'я (ВООЗ) по ліквідації віспи.
Успіхи активної імунізації (вакцинопрофілактики) дозволили ВООЗ прийняти в 1974 році "Розширену програму імунізації", яка передбачає проведення масових щеплень дітям в усіх країнах світової спільноти проти шести основних інфекційних хвороб: туберкульозу, поліомієліту, ко¬люшу, діфтерії, правця і кору.