

Образование белковых молекул и взаимосвязь азотного и углеродного метаболизма
Известно, что белковые молекулы состоят из аминокислот. Аминокислоты соединены связями, богатыми энергией. Белок является высоко энергетической «валютой» клетки в связи с тем, что полипептидные связи очень энергоемки. Аминокислоты не проявляют кислотных свойств вследствие наличия в одной и той же молекуле аминокислоты как карбоксильной, так и аминной группы:
H- N-C-C-C-C=O
 H
OH
H
OH
Но самым важным следствием их строения является наличие в их составе двух элементов — углерода и азота. Следовательно, аминокислоты являются тем ключевым звеном, в котором происходит взаимосвязь между азотным и углеродным метаболизмом.
 Образование
белка происходит не только на свету, но
и в темноте, хотя на свету этот процесс
происходит намного интенсивнее. Поэтому
принято считать, что в клетках происходит
образование белка двумя путями: темновой
синтез белка за счет энергии дыхательного
метаболизма и световой (первичный)
синтез белка в хлоропластах за счет
энергии фотосинтеза (Рис. 38).
Образование
белка происходит не только на свету, но
и в темноте, хотя на свету этот процесс
происходит намного интенсивнее. Поэтому
принято считать, что в клетках происходит
образование белка двумя путями: темновой
синтез белка за счет энергии дыхательного
метаболизма и световой (первичный)
синтез белка в хлоропластах за счет
энергии фотосинтеза (Рис. 38).


Рис. 38. Пространственное разделение синтеза белка в растениях.
Было установлено, что в образовании белков в хлоропласте используется азот листа. Образование белковой молекулы : как азотистой ее части, так и части, органической природы (содержащей углерод) происходит в процессе фотосинтеза и неразрывно связано и определяется доступностью азота и углерода, производимого в процессе фотосинтеза. Итак, белок является непосредственным продуктом фотосинтеза. Он образуется в хлоропластах только тогда, когда в них идет процесс фотосинтеза. Однако, сборка аминокислотных молекул осуществляется на основе более «ранних» продуктов фотосинтеза, чем углеводы (Андреева, 1965). Белок является одним их конечных продуктов метаболизма растения. Его образование, как уже говорилось, определяется доступностью фотосинтетической энергии.
Перечислим факторы, оказывающие влияние на синтез белка:
-
доступность энергии ( на свету-энергии фотосинтеза)
-
уровень освещенности,
-
факторы, определяющие конкуренцию за энергию;
-
доступность углеводного субстрата
-
интенсивность фотосинтеза
-
активность трансмембранного переноса ассимилятов;
-
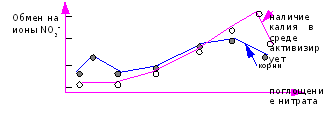
доступность азотного субстрата
-
поглощение нитратов и ионов аммония,
-
активность транспорта к листьям,
-
интенсивность восстановления нитратов.
Внешние условия и, в частности, концентрация СО2 в атмосфере могут оказывать существенное влияние на эти процессы.
Изменение азотного метаболизма при повышении концентрации со2
К настоящему времени роль минерального (и в частности азотного) питания изучена на многих культурах. Обычно исследователями ставится вопрос «какой уровень минерального питания необходим при повышенной концентрации СО2 в воздухе?».
Часть исследователей считает, что более высокая масса растений, при подкормках СО2 может быть получена только на высоком уровне минерального питания и хорошем поливе. Характерны данные, полученные на культуре перца, наглядно демонстрирующие особенность влияния повышенной концентрации СО2, вызывающего более раннее цветение на высоком уровне азотного питания (Penuelas,1995) (Рис. 39).
25 30 35 40 45 50 дни
Рис. 39. Динамика цветения у растений перца при двух режимах подкормки углекислым газом
Сегодня ситуация исследований в этом вопросе такова, что существуют доказательства двух противоположных гипотез (Рис. 40):
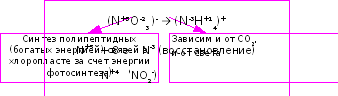
Рис. 40. Гипотезы изменения содержания концентрации азота в растениях при подкормках СО2 (Penuelas et al., 1995)
Причина этого может заключаться в интенсивном росте таких растений (обогащаемых СО2) на фоне снижения транспирации (при повышенном уровне СО2 транспирация снижается на 20-30%), что уменьшает интенсивность передвижения питательных веществ по ксилеме. Любые сообщения о влиянии высокого и низкого уровня азота на рост и развитие растений при подкормках СО2 следует воспринимать критически с той точки зрения, что в любом изучаемом диапазоне доз есть понятие оптимума (Рис 41): Поставим несколько точек на представленной выше кривой: 1,2,3,4,5,6. В зависимости от того, в какой диапазон «оптимума» мы при этом попали, мы получим тот или иной эффект от СО2.
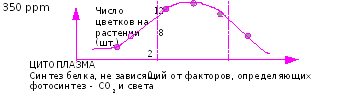
Рис. 42. Положение «оптимума» в реакции растений на увеличение доз азотного питания.
Приведем «гипотетический» график эффекта от обогащения углекислым газом в зависимости от уровня минерального питания (Рис.43).
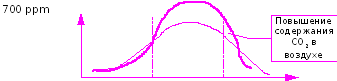
Рис. 43. Эффект от подкормок СО2 в зависимости от уровня азотного питания
Приведенный рисунок показывает, что при использовании углекислотных подкормок, чрезвычайно важно иметь представление о том, какой диапазон в указанной системе «оптимума» минерального питания мы захватили. Ожидать значительного эффекта от обогащения воздуха СО2 при неблагоприятном режиме минерального питания бессмысленно.
Если авторы исследований говорят о том, что на высоком уровне питания (точка 2 на рис.42) не была получена большая продуктивность, чем на низком (точка 1), то становиться понятным, что выбранный «высокий» уровень является высоким только по отношению к точке «1», но не является оптимально высоким, необходимым для получения эффекта от СО2.
Какие же дозы минерального питания предпочтительны и что считать нормой при обогащении растений СО2?
Исследования показали, что оптимально высокие дозы питания, дающие максимальную продуктивность растений при естественной концентрации СО2 в воздухе, будут являться оптимальными и при повышенной концентрации СО2. Однако диапазон оптимума будет уже (Рис. 44).
Случайный выбор уровня питания может оказаться основным фактором, не позволяющим гарантировать высокий уровень продуктивности при подкормках углекислым газом.
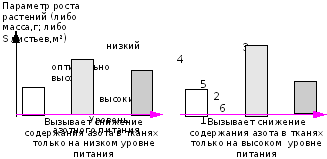 (г.
на растение)
(г.
на растение)
Уровень минерального питания
Рис. 44. Влияние уровня минерального питания на эффект от обогащения СО2 (эффект- продуктивность растений)
При выращивании растений в условиях гидропоники кажется, что растения обеспечены минеральным питанием на самом высоком уровне. Однако, здесь важно учитывать, что подача питательного раствора определенной концентрации, обеспечивающей «оптимально высокий» уровень питания, осуществляется, например, один -два раза в неделю(Рис 45.).

Рис.45. Изменение концентрации питательного раствора при выращивании растений в условиях гидропоники.
Когда растения маленькие, им хватает данной концентрации питательных веществ в режиме «оптимальный уровень» надолго. Но растения растут, и поэтому, концентрация питательного раствора снижается. Причем, снижается она значительно, часто, начиная уже со следующего дня роста (после подачи раствора) Получается, что растения (особенно взрослые), ежедневно «выедая» из раствора питательные вещества, растут при постоянном «недостатке» питательных веществ, и только раз в неделю для их роста обеспечивается «оптимум» питания
Вспомним начало главы, где указывалось, что при резком изменении концентрации питательного раствора поглощение томатом азота происходит большей частью в аммонийной форме (на фоне которой ассимиляция СО2 меньше). В результате, даже в момент смены раствора (когда минеральное питание действительно «оптимально»), растения не могут ассимилировать СО2 со скоростью, необходимой для максимальной эффективности использования питательного раствора.
Все вышесказанное ни в коем случае не говорит о том, что повысить продуктивность растений обогащением СО2 воздуха невозможно. Однако, следует понимать, что это не столь легко и существует много причин, ограничивающих применение этого метода. Выбор оптимального режима возможен при исследовательском подходе к ситуации. Например, более частая смена раствора (пусть даже чуть меньшей концентрации) может быть осуществлена параллельно в одной из обогащаемых СО2 теплиц.
ВОПРОСЫ:
-
Какая из цепочек превращения азота реально существует в растениях:
-
NО3- à NO2- à NH4+
-
NО2- à NO3- à NH4+
-
NH4+ à NО2- à NO3-
-
NH4+ à NO3-
-
Какая из форм азотного питания (NH4+ или NO3-) требует дополнительных затрат энергии для ассимиляции ?
-
От каких факторов может зависеть ассимиляция нитрата?
-
Сколько активных центров имеет нитратредуктаза, и что означает соотношение 1:3 применительно к этому ферменту?
-
5. Почему растения могут испытывать недостаток при постоянной (раз в неделю) подаче питательного раствора «оптимальной» концентрации
Ключевые слова:
нитратредуктаза, оптимум минерального питания, содержание азота в тканях.