

Снижение активности работы рдФк
Мы уже говорили о том, что за периодом активации работы РДФк при повышении СО2 в воздухе следует период снижения ее активности. Причины этого пока неизвестны. Однако, идет разработка различных гипотез тех механизмов, которые это обеспечивают. Существуют данные о снижении в атмосфере, обогащенной СО2 количества фермента РДФк (Leegood,1988).
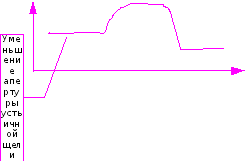
Интенсивная карбоксилирующая активность этого фермента с течением времени падает, причем активность может снижаться даже ниже его активности при низкой концентрации СО2(Рис.28).
В первое время, фиксируя неорганические молекулы СО2, РДФк активно производит ФГК (первый продукт карбоксилирования) — скорость синтеза возрастает в 12.5 раз. ФГК является специфическим активатором реакций биосинтеза крахмала. Следовательно, накоплению именно этого продукта в клетках растения будет особенно способствовать обогащение атмосферы СО2.
активность РДФк
350ppm высокая концентрация СО2
Рис. 28. Активность фермента РДФк при естественной концентрации СО2 в воздухе (1) и при повышенной концентрации СО2. (2)
В опытах Мокроносова (1981), при повышении концентрации СО2 до 1000ppm и увеличении интенсивности фотосинтеза в 3.8 раз скорость синтеза крахмала возросла в 10.7 раз. Соответственно активизации фотосинтеза увеличился и синтез сахарозы — в 4 раза.
На рисунке 29 изображена локализация синтеза крахмала и сахарозы.
Рис. 29. Активация производства углеводов — крахмала и сахарозы.
Видно, что синтез крахмала происходит в хлоропласте и при увеличении интенсивности фотосинтеза, вызванного активизацией поглощения СО2 из воздуха, накопление крахмала в клетке является прямой ответной реакцией.
Синтез сахарозы, в отличие от крахмала, локализован в цитоплазме и зависит от двух процессов:
-от активности карбоксилирования (интенсивность фотосинтеза);
-от активности трансмембранного переноса метаболитов и АТФ.
Таким образом, синтез сахарозы зависит не только от того, с какой скоростью образуются метаболиты, но и от того насколько доступна энергия и насколько активно работают переносчики метаболитов через мембрану хлоропласта.
Итак, при повышении концентрации СО2 специфичным оказывается резкое увеличение синтеза крахмала. Многие исследователи считают, что избыточное накопление крахмала в листьях является причиной снижения интенсивности фотосинтеза в атмосфере с высоким содержанием углекислого газа. Однако механизм этого явления до сих пор детально не раскрыт. Считают, что крахмальные граны механически негативно воздействуют на тиллакоиды, либо просто уменьшают свободное пространство стромы хлоропласта. Кроме того, возможно торможение выработки ФГК по принципу обратной связи: накапливающееся количество крахмала приводит к появлению «сигнала» по уменьшению производства субстрата для крахмала — ФГК, а следовательно и поглощения СО2.
В продолжение этой гипотезы рассмотрим возможность ингибирования фотосинтеза по принципу «обратной связи», то есть, снижения производства углеводов из-за накопления их в органах. Этот механизм эндогенной регуляции мы рассмотрим отдельно.
Кроме того, при высоком уровне СО2 неэффективный фотосинтез, не приводящий к увеличению накопления большей массы растений, может быть следствием неоптимального соотношения дыхания и фотосинтеза. Растения могут просто «сдышать» всю биомассу.