

Перенос ассимилятов на дальние расстояния
Передвижение производимых в процессе фотосинтеза ассимилятов на дальние расстояния — к тканям нефотосинтезирующим осуществляется посредством передвижения по ситовидным клеткам флоэмы. Поэтому, чтобы оценить активность передвижения веществ по флоэме, следует определить ее «производительность». Флоэма является не просто проводящим путем между акцептором (потребляющими тканями) и донором (производящими ассимиляты тканями), но проводящей системой с целым рядом условий, которые необходимы для поддержания функций этой системы. Флоэма состоит из проводящих пучков, в которые собраны ситовидные трубки, осуществляющие транспорт. Для проводящей системы характерны очень высокие величины дыхания: дыхание таких клеток в несколько раз превосходит дыхание клеток окружающей паренхимы. Многие проводящие клетки флоэмы лишены своего содержимого, но «клетки-спутницы», входящие в комплекс проводящей системы, характеризуются повышенными величинами дыхания.
Можно обратить особое внимание на существенную роль возможного подавления дыхания целого растения (различными внешними факторами) в изменении транспорта,а следовательно, и фотосинтеза. Поскольку известно, что посредством дыхания у проводящих элементов поддерживается резерв энергии, необходимый для транспорта по ним веществ. Составим логическую схему, связывающую эти процессы (Рис. 32).
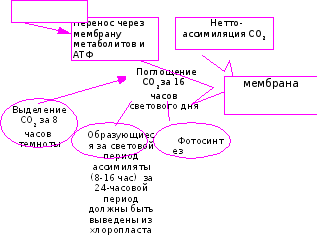
Рис 32. Логическая схема зависимости транспорта веществ по флоэме от различных факторов
Транспорт веществ по ситовидным трубкам может резко снижаться при охлаждении, однако следует отметить, что его температурные границы очень широки. Значительное повышение температуры также может ослаблять транспорт, но здесь избыточные затраты на дыхание ограничивают доступность энергии для транспорта.
Транспорт по флоэме зависит от света. Это определяется на первом этапе загрузкой флоэмных окончаний ассимилятами, а также энергией, необходимой для этого. Эти два параметра зависят от света в той мере, в которой зависит от него фотосинтез. Однако, интенсивность света в обычных условиях не влияет на скорость передвижения веществ во флоэме непосредственно. То есть, влияние света скорее опосредованное — через интенсивность работы ферментов и механизмов, определяющих «загрузку» и «разгрузку» флоэмы. В световой период энергия, необходимая для «загрузки» ассимилятами флоэмы, черпается из фотосинтетических и дыхательных резервов АТФ, а в темновой период — только из дыхательных резервов. По этой причине ночная активность передвижения веществ может быть менее интенсивной.
Исследования передвижения по флоэме сахарозы как основной транспортной формы углеводов показали, что она передвигается по флоэме «порциями» по градиенту концентраций (Рис. 33):
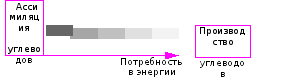
Рис. 33. Движение сахарозы по градиенту концентраций между листом (донор) и корнем (акцептор).
В связи с этим становится понятен механизм, называемый механизмом «обратной связи», когда производство углеводов начинает ограничиваться потреблением: в первую очередь происходит это на стадии выравнивания градиентов концентраций, наличие которых в большой мере определяет передвижение веществ.