


Регуляция обмена гликогена 53
каскад, приводящий к фосфорилированию (глюкагон и адреналин) и дефосфорилированию ферментов.
А Аллостерическая регуляция
Ферменты гликогенсинтаза и гликогенфосфорилаза способны «реагировать» на уровень метаболитов и энергии в клетке. Синтез гликогена — гликогеногенез — активен при большом количестве субстратов и энергии в клетке. Распад гликогена — гликогенолиз — происходит, когда количество глюкозы и энергии снижается.
Прежде чем приступить к описанию механизма аллостерической регуляции, рассмотрим сами ферменты, которые ей подвержены.
Гликогенфосфорилаза
Гликогенфосфорилаза катализирует отщепление остатков глюкозы от молекулы гликогена. Этот фермент является аллостерическим и может находиться может находиться в двух состояниях:
1.Фосфорилированное состояние является активным и называется a-формой. Стимулируется глюкагоном и адреналином. Особенностью данного состояния является его невосприимчивость к аллостерическим ингибиторам (глюкозо-6-фосфату, АТФ).
2.Дефосфорилированное состояние является неактивным и называется
b-формой. Стимулируется гормоном инсулином.
Каждое из этих состояний может принимать одну из двух конформаций (см.
Рис. 17):
1.R-конформация активна и хорошо связывает остатки фосфорной кислоты (субстрат реакции). Превалирует у фосфорилированных молекул фермента.
2.T-конформация неактивна и не способна связываться с остатками
фосфорной кислоты. Преобладает у нефосфорилированной гликогенфосфорилазы.
Фосфорилирование и дефосфорилирование гликогенфосфорилазы происходит под действием гормонов и относится к гормональной регуляции. Аллостерическая регуляция позволяет изменять конформацию фермента: T ↔ R.
Поведение гликогенфосфорилазы соответствует параллельной модели кооперативного взаимодействия: связывание аллостерического эффектора одновременно меняет конформацию обеих субъединиц фермента.
Между мышечной и печёночной гликогенфосфорилазой есть небольшие отличия в аллостерической регуляции:
1.Печёночная гликогенфосфорилаза ингибируется глюкозо-6-фосфа-
том, АТФ и глюкозой (т.е. переходит в неактивную T-конформацию). Как правило, в печени фермент обычно находится в активной a-форме. При избытке глюкозы он переходит из R-конформации в неактивную T-конформацию.
2.Мышечная гликогенфосфорилаза ингибируется глюкозо-6-фосфа-
том и АТФ (переходит в Т-конформацию) и активируется АМФ (принимает активную R-конформацию). При больших физических нагрузках в

54 |
Глава 3 |
Обмен гликогена |
мышцах образуется много молекул АМФ, которые сигнализируют о резком снижении «доступной энергии» и необходимости в глюкозе для работы мышц. В расслабленных мышцах гликогенфосфорилаза чаще всего находится в неактивной b-форме. Как только возникает потребность в глюкозе, АМФ способен активировать даже неактивную гликогенфосфорилазу b, изменив её конформацию с T на R.
Рис. 17. Регуляция гликогенфосфорилазы. Фермент может находится в 4 состоя-
ниях: фосфорилированном T и R и дефосфорилированном T и R. Фосфорилированное состояние (а-форма) является активным для фосфорилазы. При этом фосфорилированная форма может иметь как R-конформация (более активную), так и T- конформация (менее активную). То же относится и к дефосфорилированной форме (b-форме). Самым активным является состояние a+R, а наименее активным — b + T (зелёным цветом отмечена самая активная форма, а красным — наименее активная).
Биохимический смысл аллостерической регуляции гликогенфосфорилазы заключается в том, что избыток продукта подавляет активность фермента. Если глюкозы и энергии в клетке стало более, чем достаточно, то совершенно ни к чему расщеплять гликоген, поэтому глюкозо-6-фосфат и АТФ подавляют гликогенфосфорилазу.
Гликогенсинтаза
Гликогенсинтаза участвует в синтезе гликогена и существует в двух формах:
1.Фосфорилированная форма является неактивной и называется гликогенсинтазой I (b-форма, глюкозо-6-фосфат независимая). Не способна

Регуляция обмена гликогена 55
к аллостерической регуляции. Стимулируется глюкагоном и адреналином.
2.Дефосфорилированная форма активна и называется гликогенсинтазой D (a-форма, глюкозо-6-фосфат зависимая). Стимулируется инсулином. Эта форма способна к аллостерической регуляции: активируется глюкозо-6-фосфатом.
БГормональная регуляция
Глюкагон и адреналин
Процессы синтеза и расщепления гликогена должны быть строго синхронизированы с состоянием организма (голодание и насыщение), поэтому гормональная регуляция обмена гликогена крайне важна.
При голодании альфа-клетки островков Лангерганса поджелудочной железы секретируют гормон глюкагон. В стрессовом состоянии и при физических нагрузках хромаффинные клетки мозгового вещества надпочечников выделяют другой гормон
— адреналин. Глюкагон и адреналин действуют на клетки печени и мышечной ткани.
Внутриклеточный сигнальный каскад, запускаемый этими гормонами, активирует распад гликогена и одновременно выключает его синтез.
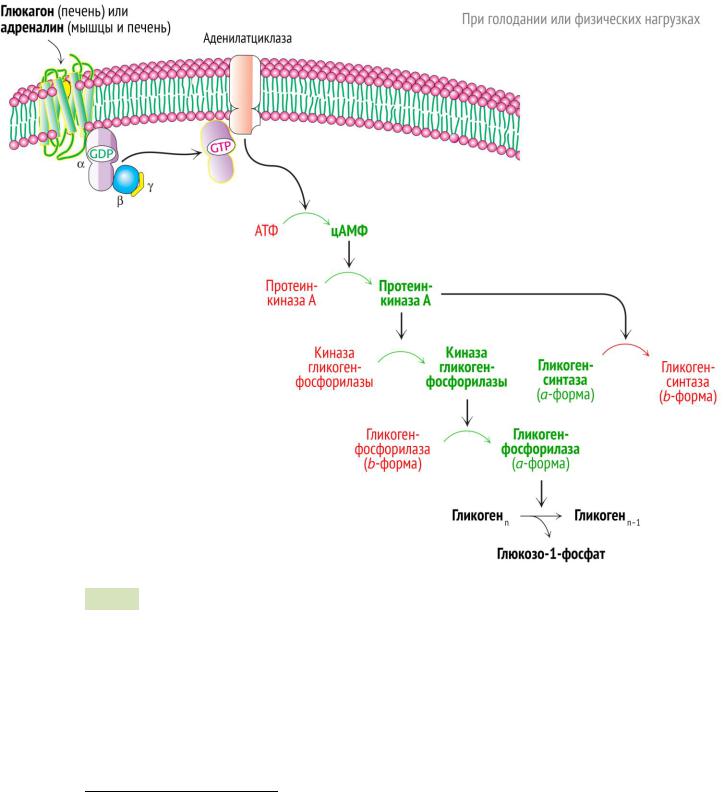
Этапы регуляции (см. Рис. 18):
1.При голодании снижается уровень глюкозы в крови, на что сразу же реагируют клетки поджелудочной железы и секретируют глюкагон. При стрессе в ответ на нервные импульсы клетки надпочечников выделяют адреналин.
2.Глюкагон и адреналин связываются со своими рецепторами на по-
верхности клеток и запускают аденилатциклазную сигнальную систему.
а. Наибольшее количество рецепторов к глюкагону находится на поверхности клеток печени, поэтому печень является основным эффекторным органом глюкагона (небольшое количество рецеп-
торов к нему обнаружено в мембранах клеток сердечной мышцы, жировой ткани, селезёнки, тимуса, поджелудочной железы, ЖКТ и коры головного мозга).
б. Адреналин связывается с β-адренергическими рецепторами клеток печени и мышц и с α1-адренергическими рецепторами клеток печени.
3.Запуск сигнальной системы приводит к активации фермента аденилатциклазы, который синтезирует вторичный мессенджер — циклический АМФ (цАМФ).
4.цАМФ активирует фермент протеинкиназу А, фосфорилирующий ферменты, участвующие в метаболизме гликогена:
а. Фосфорилированная гликогенсинтаза переходит в неактивную b-форму. Синтез гликогена прекращается.
56 |
Глава 3 |
Обмен гликогена |
|
б. |
Фосфорилированная киназа гликогенфосфорилазы1 (активная) |
|
|
фосфорилирует гликогенфосфорилазу и переводит её в актив- |
|
|
ную a-форму. |
5. Гликогенфосфорилаза начинает расщеплять гликоген. Таким образом, запускается распад гликогена в печени и мышцах.
Рис. 18. Регуляция расщепления гликогена под действием гормонов глюкагона и адреналина.
Инсулин
Инсулин — анаболический белковый гормон, активирующий синтез гликогена в печени и мышцах. В течение первого часа после приёма богатой углеводами пищи концентрация глюкозы в крови повышается до 150 мг/дл, а затем снижается до нормальных значений в течение 2-3 часов.
1Киназа гликогенфосфорилазы — регуляторный фермент, фосфорилирующий гликогенфосфорилазу (и активирующий её).

Регуляция обмена гликогена 57
Клетки поджелудочной железы очень чувствительны к повышению сахара в крови, поэтому секретируют инсулин в кровь, как только количество глюкозы в крови резко возрастает.
Инсулин понижает концентрацию глюкозы в крови, вызывая целый ряд событий и стимулируя захват глюкозы клетками и синтез гликогена в печени и мышцах. Инсулин-
зависимые ткани (мышечная и жировая) захватывают глюкозу благодаря транслокации глюкозных транспортеров (ГЛЮТ4-белков) из цитозоля в мембрану клеток. Попав в клетку, глюкоза фосфорилируется до глюкозо-6-фосфата, который аллостерически активирует фермент гликогенсинтазу, переводя его из неактивной T- конформации в активную R-конформацию (даже если он находится в b-форме).
Рис. 19. Регуляция биосинтеза гликогена под действием гормона инсулина.
Этапы регуляции:
1.После приёма пищи, богатой углеводами, уровень сахара в крови резко повышается, и бета-клетки поджелудочной железы начинают секрети-
ровать инсулин.
2.Инсулин связывается со своим рецептором в мембране клетки (печени или мышечной ткани).
3.Активируется тирозинкиназная активность рецептора, и он фосфори-
лирует особый белок — субстрат инсулинового рецептора (IRS-бе- лок).