

Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
136 |
Пол у E.coli
В 1946 году Джошуа Ледерберг и Эдвард Тэйтем принялись ставить генетические эксперименты на бактериях. Несколькими годами ранее Тэйтем работал в сотрудничестве с Джорджом Биллом, и они на основе опытов с плесенью Neurospora выдвинули теорию «один ген — один фермент», оказавшую столь большое влияние на развитие генетики. Ледерберг и Тэйтем надеялись при помощи более простых организмов сделать еще более грандиозное открытие. Лурия и Дельбрюк к тому времени уже доказали, что бактерии подобно другим организмам могут мутировать и образовывать разные штаммы, полезные в исследованиях. Однако для генетических экспериментов требуется скрещивать различные линии организмов с половым размножением, тогда как у бактерий, как считали в то время, пола нет. Ледерберг и Тэйтем предположили, что если бы у бактерий имелся пол, то разные клетки с разным генотипом скрещивались бы между собой и давали рекомбинантов, как и более высокоразвитые организмы. Поэтому если обнаружить рекомбинантные организмы, то это будет доказательством того, что у бактерий имеется нечто вроде полового размножения. Они предположили, что даже в этом случае количество рекомбинантов будет очень малым, и поэтому нужно постараться вывести рекомбинан-
253
тов между двумя ауксотрофными штаммами. Один из штаммов для своего роста требует треонин и лейцин, но может производить свои метионин и тиамин (генотип thr-leu-met+thi+); другой штамм может вырабатывать треонин и лейцин, но требует метионин и тиамин (генотип thr+leu+met-thi-). При смешении этих штаммов действительно обнаруживались прототрофы, которые могли образоваться только в результате рекомбинации.
Леденберг обнаружил, что рекомбинации могут происходить только между некоторыми фертильными штаммами, которые он обозначил F+, и другими штаммами с довольно низкой частотой, которые он обозначил F", притом с довольно низкой частотой. Фертильность у бактерий — это нечто вроде инфекции, потому что когда клетки F- смешиваются с клетками F+, они преобразуются в клетки F+. Такое странное явление прояснили эксперименты Уиляма Хэйза из Великобритании, Франсуа Жакоба и Эли Вуллмана из Франции и Л. Л. Кавалли из Италии. Сначала они открыли, что клетки F+ содержат генетический фактор, названный ими F-фактором, или фертильным фактором. Как мы увидим далее, F-фактор представляет собой отдельную молекулу ДНК. При контакте с другими клетками клетки F+ способны передавать копии своих F-факторов другим клеткам, преобразуя их в клетки типа F+. С низкой вероятностью они могут заодно передавать и некоторые свои гены клеткам F-, и так происходят рекомбинации. Затем Хэйз и Кавалли открыли вариант штамма F+, названный Hfr (high frequency of recombination — «высокая частота рекомбинации»), который передает свои гены клеткам F" с довольно высокой частотой.
254
Механизм рекомбинации стал ясен после скрещиваний прототрофного штамма Hfr, чувствительного к стрептомицину, со множественным ауксотрофным штаммом F-, устойчивым к антибиотику, и с генотипом strr thr-leu-met-lac-gal-thi-. Каждый ген означает метаболический недостаток, например неспособность синтезировать треонин (thr~) или неспособность
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.
Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
137 |
расти на лактозе (1ас-). При смешивании клетки Hfr и F- спариваются в процессе конъюгации (слияния). Через некоторое время их помещают в среду со стрептомицином, чтобы убить клетки Hfr и проверить, какие дикие гены клетки F" унаследовали от клеток Hfr. В ставшем классическом эксперименте Жакоб и Вуллман через разные промежутки времени брали образцы конъюгирующих клеток и разделяли их в смесителе, после чего выращивали в различных средах и определяли унаследованные гены. Этот эксперимент показал, что каждый маркер Hfr начинает проявляться через строго определенный промежуток времени: ген В только через семь минут, ген С — через девять, ген D — только через 15 минут и т. д. Отсюда следует, что Hfr передает клеткам F- гены в линейной последовательности, то есть в той последовательности, в которой они соединены в хромосоме бактерий.
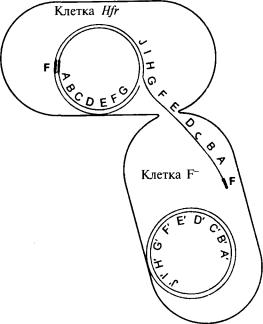
На рис. 10.1 объясняются полученные результаты. Фактор F в клетках F+ представляет собой небольшую кольцевую молекулу ДНК. Контакт с клетками F- побуждает ее к быстрой репликации и переносу реплики в клетки F-, благодаря чему они превращаются в клетки F+. Во время этого процесса некоторые гены F+ случайным образом проникают в клетку F-, хотя до сих пор не совсем ясно, как это происходит. Как бы то ни было, в клетках
Hfr
255
Рис. 10.1. Перенос ДНК при скрещивании Hfr х F.
Клетка Hfr соединяется с клеткой F-, и между ними образуется конъюгационная трубка. Фактор F, встроенный в хромосому Hfr, начинает реплицироваться. Но так как фактор F встроен в хромосому, он заодно запускает репликацию всей хромосомной ДНК. Поэтому в клетку F- переходит копия хромосомы Hfr с генами в линейной последовательности. Как только донорская ДНК проникает в клетку F-, она может спариваться с хромосомой F- и образовывать рекомбинанты с аллелями Hfr в хромосоме F-
фактор F встраивается в хромосому бактерии, которая тоже кольцевая, хотя и больше по размеру. Теперь при контакте с F" фактор F снова начинает переносить свою копию, но поскольку фактор F и
256
бактериальная хромосома представляют собой единое целое, эта копия
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.
Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
138 |
содержит бактериальные гены, которые передаются последовательно: А, В, С, D и т. д. Конъюгация редко длится так долго (90-100 минут), чтобы произошла репликация всей кольцевой хромосомы вплоть до начала фактора F, поэтому клетки F-, как правило, не так уж часто преобразуются в клетки Hfr.
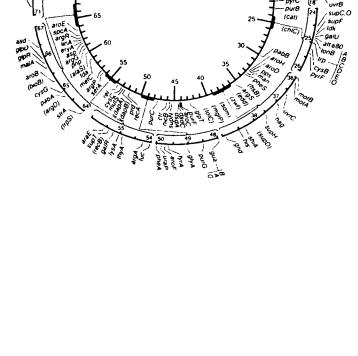
После конъюгации вновь перенесенная ДНК Hfr рекомбинирует с хромосомой F-, и появляются разнообразные рекомбинанты. Но в данном случае по частоте рекомбинации определяют не общую карту Е. coli и расстояние между генами в единицах, а расстояние в минутах между маркерами. Фактор F может встраиваться в различные места и переноситься в разных направлениях от этой точки; каждое такое встраивание приводит к образованию отдельного штамма Hfr уникального происхождения. Так строят карту различных участков генома. Подробные генетические карты Е. coli (рис. 10.2) и других бактерий были составлены при исследовании всех этих штаммов, а также при помощи других методов, которые будут описаны далее.
Плазмиды
Фактор F — пример так называемой плазмиды, то есть внехромосомного самореплицирующегося генетического элемента с кольцевой структурой. Плазмиды — это своего рода пассажиры в клетке, которые реплицируются при репликации клеточной ДНК. Так как они сами представляют собой часть молекулы ДНК, то в них, как и в вирусах, могут содер-
257
Рис. 10.2. Карта хромосомы Е. coli.
Единицами служат минуты, то есть промежутки времени, через которые переносятся гены при скрещивании Hfr х F-. Отдельные участки карты с большим количеством известных генов более подробно показаны на внешних сегментах. Более мелкие подробности определяются в процессе трансдукции, о чем говорится далее. Данная карта была опубликована несколько лет назад, и сейчас существуют более подробные карты, которые заняли бы несколько страниц
жаться различные гены, однако в отличие от вирусов плазмиды не образуют внеклеточных частиц (вирионов) ни на одной из стадий своего
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.