

Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
117 |
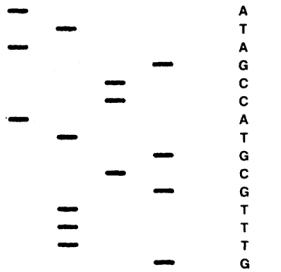
Рис. 8.4. Один из методов определения последовательности молекулы ДНК
В четырех пробирках синтезируются цепи ДНК, комплементарные очищенным одиночным цепям, причем в каждой пробирке содержатся различные дидезоксинуклеотиды. Получившиеся фрагменты распределяют в зависимости от размера и всю по-
218
следовательность прочитывают по распределению полос.
Рестрикционное картирование
Сейчас известно и доступно для применения множество типов рестрикционных ферментов. Они разрезают ДНК на различные последовательности, и их можно использовать для анализа структуры ДНК и составления хромосомных карт методом рестрикционного картирования. На рестрикционной карте отмечено относительное расположение участков (сайтов рестрикции), которые вырезают различные нуклеазы. С помощью других методов можно сопоставить эту карту с генетической картой. В данном случае применяют технологию электрофореза, описанную во вставке (с. 216-218), с помощью которой разделяют ДНК на фрагменты и определяют их относительный размер. Рестрикционное картирование лучше объяснить на примере. У многих вирусов животных имеются маленькие кольцевые ДНК. Предположим, мы выделили ДНК вируса длиной в четыре килобазы (kb) и порезали ее ферментом ЕсоRI на фрагменты. Пропустив их через гель, мы определили их длину: 0,4; 0,8; 1,3 и 1,5 kb. Это значит, что в геноме находится четыре участка рестрикции EcoRI, которые могут располагаться по-разному.
Порежем вирусную ДНК снова при помощи EcoRI, но на этот раз уменьшим время обработки ДНК ферментом, чтобы некоторые ДНК были порезаны не полностью. Наряду с прежними четырьмя фрагментами получаем новые фрагменты длиной 1,7;
219
1,9; 2,1 и 2,3 kb. Небольшой перебор вариантов показывает, что эти фрагменты располагаются в следующем порядке:
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.

Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
118 |
Можно подтвердить такое расположение, выделив более крупные фрагменты, порезав их ЕсоRI и убедившись, что они в итоге разрезаются на те же малые фрагменты.
Далее обработаем ту же вирусную ДНК ферментом Sali, в результате чего получим фрагменты длиной 0,95; 1,25 и 1,8 kb. Получается, что в этой ДНК три участка SalI. Затем выделим эти фрагменты и порежем их ЕсоRI:
фрагмент 0,95 kb -> 0,1 kb + 0,85 kb; фрагмент 1,25 kb -> 0,2 kb + 0,4 kb + 0,65 kb; фрагмент 1,8 kb -> 0,7 kb + 1,1 kb.
Перебор вариантов показывает, что участки SalI располагаются относительно участков ЕсоRI следующим образом:
220
Если полученных данных недостаточно для определения взаимного расположения участков, то можно еще раз порезать фрагменты одним ферментом, а затем другим, или, допустим, порезать ДНК при помощи сначала EcoRI, а затем SalI. После этого можно применить третий фермент и определить расположение его участков рестрикции относительно первых двух. Это очень простой пример. На практике дело обстоит гораздо сложнее даже в случае с самыми маленькими вирусами, которые требуют более сложных схем анализа, но принцип анализа сохраняется.
Эту же технологию можно применять для диагностики наследственных нарушений по ДНК зародышевых клеток. Обнаружилось, что фермент Hpal режет нормальную ДНК на фрагменты одной длины, а ДНК с аллелями серповидноклеточной анемии гена бета-гемоглобина — на фрагменты другой длины; в 87% случаев серповидноклеточной анемии получаются более длинные фрагменты. Это
221
значит, что дефектный участок гена с мутацией HbS не соответствует последовательности фермента Hpal. При помощи этого метода возможно определять 87% случаев серповидноклеточной анемии непосредственно на стадии эмбрионального развития.
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.

Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
119 |
Рестриктазы используют и в других анализах. Во всех биологических видах наблюдается некоторое разнообразие в последовательности ДНК различных особей, и иногда такое разнообразие приводит к удалению или вставке дополнительных участков рестрикции. Обычно это нейтральные вариации, не оказывающие влияния на фенотип (в отличие от вариации гена гемоглобина). Такие вариантные участки оказываются весьма полезными при составлении карт, потому что они говорят об альтернативной форме хромосомы с наличием или отсутствием дополнительного участка рестрикции:
или:
Молекулярный анализ легко обнаружит эти различия, потому что если эту последовательность ДНК разных индивидов порезать рестриктазой, то в одном случае получится один длинный фрагмент, а в другом — два коротких, общая длина которых соответствует длине первого. Таким образом выявляются два морфа в популяции: у некоторых индивидов имеется дополнительный участок рестрикции, а у других его нет. Такое явление называется полиморфизмом длины рестрикционных фрагментов
(RFLP— restriction fragment length polymorphism). Его полезно учитывать при составлении карт, потому что этот
222
полиморфизм служит нейтральным гетерозиготным маркером, с помощью которого можно определять близлежащие гены, особенно при составлении карт маркеров, приводящих к разным фенотипам. Кроме того, если RFLP расположен близко от дефектного аллеля, то его можно использовать для обнаружения этого аллеля подобно участку рестрикции внутри гена гемоглобина. В геноме встречаются и другие типы нейтральных вариаций последовательности, и RFLP оказался первым среди открытых учеными. Все они могут быть использованы для составления карт и обнаружения рецессивных дефектных аллелей.
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — М.: ФАИР-
ПРЕСС, 2004. — 448 с: ил.