

1 СЕМЕСТР. Экономика. Микроэкономика. Поведение, институты и эволюция Самуэль Боулз / Микроэкономика. Поведение, институты и эволюция_Самуэль Боулз_2010 -576с
.pdfГлава 13. Совместная эволюция институтов и предпочтений 439
Таблица 13.4
Оценка уравнения Прайса
Институты |
Коэффициент влияния |
Отношение дисперсий |
p |
|
|
|
|
Ни одного |
0,25 |
0,13 |
0,06 |
|
|
|
|
Оба |
0,13 |
0,13 |
0,51 |
Примечание. Отношение дисперсий — это среднее значение var (pj)/E {var (pij)} по 10 тыс. поколений, а отношение влияний -βi /βG оценивается так, как описано в табл. 13.2. Средняя доля представителей типа А в популяции равна p.
Конфликты между группами играют ключевую роль в поддержании инсти тутов на уровне групп и альтруизма на уровне индивидов. В построенной модели ожидаемая частота возникновения конфликтов составляла 1/k, где k — вероят ность того, что в каждом поколении группу выберут для соревнования. Кажется вероятным, что на протяжении большого числа исторических периодов частота возникновения конфликтов сильно изменялась, возможно, в ответ на необхо димость миграции в периоды климатических изменений. Чтобы исследовать чувствительность построенной модели к частоте возникновения конфликтов, мы изменяли k случайным образом, используя авторегрессию, описанную в при мечании к рис. 13.6. В течение периодов, когда конфликты возникали часто (т. е. примерно в районе 21 000го поколения), поддерживались высокие уровни аль труизма, но периодические случаи наступления перемирия (в районе 25 300, 27 000 и 29 600го поколений) приводили к резкому сокращению доли инди видов типа A в популяции. Период с 500 до 28 500го поколения демонстрирует зависимость от траектории предшествующего развития в этой модели. Высокий
Рис. 13.6. Высокие частоты групповых конфликтов способствуют альтруизму. Рисунок отражает период в тысячу поколений в модели, в которой оба института эндогенно развиваются и k, частота межгруппового конфликта, изменяется во времени согласно kt = k0 + ρkt − 1 + σt, где ρ = 0,99, σt равномерно распределено на [−0,02, 0,02] и k0 выбирается так, что среднее kt равно k, здесь 0,25

440 Часть III. изменения: совместная эволюция институтов и предпочтений
уровень p, вызванный резким ростом частоты появления конфликтов между группами примерно во время жизни 28 500го поколения, сохраняется, даже по мере того как при жизни последующих поколений резко снижается частота появления конфликтов. Однако «эффект ловушки» не остается постоянным: ког да k на протяжении нескольких периодов остается на уровне ниже 0,2, p резко падает.
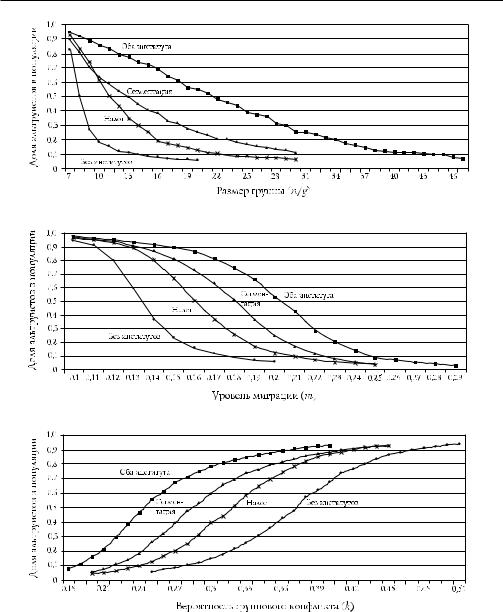
Мы также пытались ответить на два других вопроса. Могло ли развитие аль труизма не способствовать совместной эволюции институтов группового уровня и альтруизма на индивидуальном уровне? И насколько чувствительной остается наша модель по отношению к изменению ключевых параметров? Для ответа мы изменяли размер группы с 7 до 47, для каждого значения строилось 10 симуля ций по 50 тыс. поколений в каждой, при этом значения остальных параметров оставались на базовом уровне. Мы производили эти изменения в предположе нии, когда развитие обоих институтов было ограничено, когда ограничивалось развитие только одного из институтов и когда развитие институтов не ограни чивалось. Мы произвели те же операции для уровня миграции от 0,1 до 0,3 и вероятности возникновения конфликта (k) от 0,18 до 0,51. Результаты показаны на рис. 13.7.
В верхней панели показано, что если действие институтов ограничено, так что ни один из них не развивается, то при размере группы, равном 7, поддержи вается высокий уровень альтруизма, но при увеличении размера до 8 доля альт руистов снижается ниже 0,3. Приняв за начальный такой размер, для которого p > 0,5, мы видим, что в отсутствие институтов критический размер равен 8, в то время как при наличии обоих институтов p > 0,5 для групп с размером меньше 22. Результаты с учетом миграции такие же. В отсутствие институтов для поддер жания p > 0,5 требуется уровень миграции 0,13 (на поколение), а при наличии обоих институтов критический уровень миграции равен 0,21. Нижняя панель показывает, что при наличии институтов возможен высокий уровень развития альтруизма при значительно меньшей частоте возникновения конфликтов меж ду группами. «Вертикальное» прочтение рисунка также имеет смысл: например, нижняя панель показывает, что для k = 0,3 p меньше чем 0,2 в отсутствие инсти тутов, но выше чем 0,8 при наличии обоих институтов1.
1 Рисунки 13.7 и 13.3 показывают, что сегрегация может оказывать более сильное воз действие, чем коллективное пользование ресурсами: сегментация сама по себе оказывает большее влияние, чем коллективное пользование ресурсами само по себе, как на ослабление направленного против А внутригруппового отбора, так и на расширение множеств значений параметров, при которых индивиды типа А составляют большую часть популяции. Это и есть результат нашего построения. Функции затрат для s и t одинаковы, но s сильнее влияет на корректировку отбора внутри группы, что видно из равенства (13.6). Пользуясь этими равен ствами, чтобы сравнить влияние s, когда t = 0, и влияние t, когда s = 0, мы видим, что первое в b/c раз больше последнего, а b > c, потому что альтруистичное поведение выгодно для группы (при нашем построении b = 2 и c = 1, так что влияние s в два раза больше влияния t). Заметим также, что из равенства (13.5) видно, что если s = c/b = 1/2, то pj = pj (1 − pj)(1 − tj)(sjb − c) = 0, но величина t, необходимая, чтобы остановить направленный против А внутригрупповой от бор, равна 1 (в случае квадратичной функции затрат, которую мы использовали, затраты на уровне группы для t = 1 в четыре раза больше затрат для s = 1/2).
Глава 13. Совместная эволюция институтов и предпочтений |
441 |
Рис. 13.7. Групповые институты увеличивают размер пространства параметров, для которого альтруистическое поведение превалирует. Каждая точка — это средняя доля альтруистов во всем населении по 10 симуляциям, 50 тыс. периодов в каждой, для параметров, отраженных на горизонтальной оси. В каждой панели остальные пара метры взяты из табл. 13.2. Каждый прогон начинался с p, t, s, равных нулю.
Линия, обозначенная «без институтов», отражает результаты, в которых t, s равны нулю; другие кривые построены в предположении, что один или два института свободно развиваются («налоги» означают коллективное пользование ресурсами). Расстояние между кривыми по горизонтали отражает расширение пространства параметров, произошедшее благодаря групповым институтам. Расстояние по вертикали показывает влияние институтов на среднее p

442Часть III. Изменения: совместная эволюция институтов и предпочтений
Вэкспериментах, когда уровень мутации варьируется с 10 −2 до 10 −5, мы получаем похожие с представленными результаты. В отсутствие институтов p остается низким, в то время как при наличии обоих институтов среднее значение p по пяти симуляциям в каждой по 100 тыс. поколений (для уровней мутаций
10−2, 10 −3, 10 −4 и 10 −5) превышает одну вторую. Среднее значение p по пяти симуляциям с уровнем мутации 10 −5 варьируется от 0,75 до 0,83; в каждом случае резкий рост p происходил между 17 150 и 25 855 поколениями, и высокие уровни p поддерживались на оставшемся отрезке эволюции. Время ожидания начала роста зависит от времени, требуемого одной группе для того, чтобы накопить значительное число альтруистов. Когда групп больше 20, время ожидания значительно сокращается. Поскольку мы положили в начале (p = 0), низкие уровни p в течение многих периодов подкрепляются очень низким уровнем миграции (меньше 10 −5).
Эволюционная среда
Мы описали процесс, в рамках которого образование таких институтов, как коллективное пользование ресурсами и сегментация, способствует возникновению среды, где распространяются выгодные для группы характеристики, и эти институты развиваются вследствие их вклада в эволюционный успех выгодных для группы характеристик. Объясняет ли эта модель процесс, в рамках которого развиваются выгодное для группы поведение и институты на уровне группы? Ответ должен зависеть от того, соответствует ли в нашем построении совместных процессов эволюции множество значений параметров реальным данным, а именно первым 50 тыс. и 100 тыс. годам существования человечества до момента значительных изменений структуры общества наряду с развитием сельского хозяйства около 11 тыс. лет назад.
О поздней эпохе плейстоцена известно мало, но мы знаем, что достаточно сложно судить о структуре организаций групп людей во время этого периода на базе современного общества (Foley, 1987; Kelly, 1995). Тем не менее с некоторой уверенностью мы можем сказать, что климат сильно менялся (Richerson, Boyd & Bettinger, 2001) и малочисленные мобильные группы сборщиков урожая, состоявшие как из родственников, так и нет, и испытывающие недостаток в общей политической структуре, были традиционной формой организации общества.
Начальный размер группы, равный 20, основывается на оценке медианного размера 235 групп охотников, установленного Бинфордом (Binford, 2001), а именно 19. Тем не менее наше приближение не совсем реалистично. Вспомним, что малый размер усиливает давление группового отбора, увеличивая межгрупповую дисперсию, возникающую, когда успешные группы удваиваются в размере и затем делятся. В реальности разделение групп происходит не вследствие случайного выбора, а в результате процесса разрешения политических конфликтов, в котором семьи и коалиции с большой вероятностью остаются вместе. Значит, разделение, вероятно, вносит вклад в увеличение дисперсии между группами,

Глава 13. Совместная эволюция институтов и предпочтений 443
неизученное в нашей модели. В исследовании о разделении народов, живущих на берегах Амазонки (Neves, 1995. P. 198), говорится:
Максимальный размер поселения ограничен степенью родства и солидарности между индивидами, происходящей из трех источников: семейные взаимоотноше ния, брачные узы и влияние политических лидеров... Разделение деревень, таким об разом, поддерживается слабостью семейных связей, вызванной ростом населения; и когда такое происходит, семьи объединяются внутри себя, но отдаляются от других семей... [Потенциальный] раскол проходит по линии разделения родословных1.
Поскольку вероятно, что существует большое число носителей выгодной
для группы характеристики и они численно и социально доминируют в группе победителе, они могут практиковать то, что Гамильтон (Hamilton, 1975. P. 137) назвал отделением подобных, сегрегируя носителей «других» характеристик до
тех пор, пока процесс распознавания черт и характеристик, коррелирующих с ними, позволяет это. Если бы это было так, то описанный ранее процесс эволю ции поддерживался бы группами гораздо большего размера.
Мало что известно о конфликтах между группами в течение ранней исто рии человечества. Мы знаем, что гибель во время войны составляет существен ную долю от общего числа смертей во многих обществах в эпоху до зарождения государственности, по данным этнографии и археологии. Среднее значение по этнографическим исследованиям обществ в эпоху до зарождения государствен ности, проведенным Киили (Keeley, 1996), равно 0,19, а по археологическим ис следованиям — 0,16. Это сравнимо с оценками на 0,1 ниже для Европы и США XX в., 0,03 XIX в. для Франции и 0,02 для Западной Европы XIX в. 200 войн между маээнга в Новой Гвинее, например, унесли 800 жизней при населении 5 тыс. человек, при этом ежегодный показатель смертности по населению (0,0032 смертей на одного человека в год) был в два раза больше, чем в Германии и Рос
сии в XX в., но сильно ниже среднего показателя в период до зарождения госу дарственности (Keeley, 1996. P. 195). Сложно сказать, были ли такие показатели
смертности привычными для поздней эпохи плейстоцена. Но, основываясь на том, что нам известно о климатических изменениях и вероятных показателях роста населения, можно порассуждать немного. Кристофер Боэм (Boehm, 2000а. Р. 19) пишет:
Разумно предположить, что плотность населения доисторического периода росла в богатой и стабильной среде, что более сплоченные и многочисленные группы начи нали конкурировать за ресурсы, и что, в конце концов, это привело бы к политиче ским конфликтам, приводящим к летальным исходам, даже если изначально ресур сов было бы достаточно… Эти различные особенности конфликтов способствовали бы периодическому увеличению силы естественного отбора, действующего на меж групповом уровне, по мере того как одни группы распадались, а другие процветали и в итоге разделялись. [В ответ на значительные климатические колебания в послед ний межледниковый период плейстоцена] группам сборщиков урожая приходилось
1 Чагнон (Chagnon, 1983. P. 141) рассматривал разделившиеся деревни Яномамо и обна ружил, что средняя степень генетического родства в деревнях, собирающихся разделиться, не ниже, чем во вновь образованных.

444 Часть III. изменения: совместная эволюция институтов и предпочтений
достаточно часто приспосабливаться, и, конечно, это часто касалось приспособления к соседним группам, как и к изменяющейся окружающей среде.
Он делает следующий вывод:
…К концу эпохи плейстоцена, когда начинало зарождаться современное человече ство, уровень вымирания групп мог резко возрасти, по мере того как бедствующие группы хорошо вооруженных охотников и незнакомцы, не обладающие четко уста новленными правилами взаимодействия на политическом уровне, часто сталкива лись, локально либо в ходе миграции на большие расстояния.
Кэрол Эмбер (Ember, 1978) собирал данные о частоте военных конфликтов среди 50 групп сборщиков урожая в прошлом и настоящем. За исключением тех, кто собирал травы и занимался оседлым сельским хозяйством, в 64% групп военные конфликты происходили каждые два года или чаще. Даже исключая тех, кто держал лошадей или занимался рыболовством (среди тех, у кого войны случались чаще), военные конфликты можно было назвать «редкими» только в 12% групп.
В то время как движение между этнолингвистическими объединениями происходило нечасто, вполне вероятно, что уровни миграции между группами, cоставлявшими эти объединения, оставались достаточно высокими. Роджерс (Rodgers, 1990) рассмотрел 13 объединений, и средний уровень миграции соста вил 22% за поколение, а максимальный (согласно Кунгу (Kung)) был меньше по ловины. Поскольку данные Роджерса относились к объединениям большего раз мера, чем группы, то эти данные могут занижать реальные уровни миграции.
Несмотря на теоретическую природу данных выводов, вероятно, что обще ственная и физическая среда поздней эпохи плейстоцена подходила под мно жество параметров, соответствующих траектории совместной эволюции, изо браженной на рис. 13.7. Если это верно, то модель многоуровневого отбора с эндогенными институтами может, по крайней мере, частично объяснить этот важный период эволюции человека.
выводы
Таким образом, видимо, отличительная предрасположенность человека к выгод ной для группы деятельности могла развиваться совместно с внедрением таких общепринятых институтов, как коллективное пользование ресурсами и соци альная сегментация. Если это верно, то такой подход может внести вклад в по нимание причин, согласно которым люди готовы делиться и кооперироваться ради достижения общих целей и в то же время готовы убивать и быть убиты ми ради абстрактных образований, называемых нациями или расами. Модель также частично объясняет, почему такое поведение встречается не столь часто у животных: многие неспособны создавать благоприятную среду, состоящую из четко определенных групп, не связанных родственными отношениями, но име ющих ясное понимание таких типов поведения, как коллективное пользование и различия «свой — чужой». И, как мы видели, без этих четко определенных структур взаимодействия эволюция типов поведения индивида, выгодных для

Глава 13. Совместная эволюция институтов и предпочтений 445
группы, маловероятна. Следующий вывод, подчеркнутый на рис. 13.6, состоит в том, что альтруизм и война развивались совместно. Ориентированное на группу поведение делает возможным выгодное для всех сотрудничество, но также дела ет возможными летальные исходы огромных масштабов, возникающие во время войны. Частые войны способны, как мы уже видели, вносить необходимый вклад в развитие альтруистических характеристик, которые как раз и способствуют началу военных действий. Для изучения динамики мы определили эндогенным образом вероятность возникновения конфликтов между группами с летальны ми исходами; предполагалось, что k изменялась совместно с долей A в популя ции. В этих симуляциях (не показаны) популяция почти все время находится в
одном из двух состояний: большие доли альтруистов и очень частые войны или мало альтруистов и нечастые войны (Bowles & Choi, 2003).
Модели многоуровневого отбора, аналогичные построенным здесь, также по казали, что наказание нарушителей норм может распространяться, когда нару
шение происходит редко и при этом наблюдается постоянно в достаточно боль ших группах. В одной из таких моделей (Boyd, Gintis, Bowles & Richerson, 2003)
используется тот факт, что затраты, направленные на наказание нарушителей норм, достаточно малы, когда нормы придерживаются почти все представители группы (это похоже на динамику модели «Каратель — Захватчик — Подельшик»
вокрестности равновесия Руссо). В данном случае относительно слабого давле ния группового отбора, возникающего изза конфликтов между группами (как
ввышеуказанной модели), достаточно для поддержания высокой частоты по
явления альтруистических карателей и высокой степени кооперации. Во второй модели с сильной реципрокностью (Bowles & Gintis, 2004) строится форма нака
зания нарушителей нормы, традиционная для охотников и сборщиков урожая: уход или изгнание из группы. В этой модели конфликты между группами и вы мирание групп не имеет значения: индивиды, подвергнутые наказанию, страда ют от ухудшения приспособления в период, когда они не состоят в этой группе. Фрирайдеры, которые придерживаются нормы, но не прибегают к наказанию, не вытесняют сильных реципрокаторов, потому что если в группе будет много фрирайдеров, то случаи нарушения нормы участятся, что приведет к сокраще нию среднего показателя приспособления группы, в которой они находятся.
В гл. 10—13 я спрашивал, как меняются институты и как совместно изменя ются люди и правила, по которым они живут. Были построены три фундамен тальных источника изменений: подстраивание к заданным извне правилам по ведения (преимущественно технологические изменения и физические условия, как в гл. 11), совместное влияние случайных событий и коллективных действий (гл. 12) и совместное влияние случайных событий и соревнования между группа ми (гл 13). В приведенных моделях предполагается, что изменения могут прийти по двум каналам, вместе с коллективными действиями и соревнованием групп соответственно, при изменении ресурсов случайным образом. В целом модели
охватывают, по крайней мере, часть проблем, подчеркнутых в начале гл. 11, а именно: важные роли, которые играют конфликт интересов, случай и коллективные действия, постоянство неэффективных институтов и крайне нере гулярные траектории изменения, что можно в целом описать как прерывистое

446 Часть III. изменения: совместная эволюция институтов и предпочтений
равновесие. Модели дают основания полагать, что хотя неэффективные институ ты могут действовать достаточно длительно, но в долгосрочной перспективе они не станут функционировать в отличие от существующих более эффективных. Мы также видели, что по двум вполне конкретным причинам (приведенным в гл. 12 и 13) в процессе развития более вероятно действие эгалитарных институ тов. В моделях, основанных на эволюционной стохастической теории игр, об ласть притяжения эгалитарных институтов шире, а в моделях многоуровневого отбора действие эгалитарных институтов препятствует неблагоприятному для альтруистов внутригрупповому отбору, что увеличивает способность группы вы живать в условиях конкуренции с другими группами. Таким образом, мы смогли
применить несколько причинных механизмов для объяснения не только инсти туционального и индивидуального развития, но и концепции эволюционных универсалий Парсонса, а именно институтов, способных возникать и распро
страняться повсеместно при многочисленных обстоятельствах, хотя изначально они были в малом количестве.
Концептуальная стратегия, лежащая в основе всех представленных здесь моделей, состояла в том, чтобы расширить несколько достаточно абстрактных подходов биологии: эволюционной стохастической теории игр и декомпозиции уравнения Прайса для процессов внутри и межгруппового отбора с помощью построения процессов взаимоотношений людей в обществе. Таким образом, эво люционный стохастический подход был расширен с учетом намеренного пре следования конфликтующих интересов посредствам коллективных действий, а модель многоуровневого отбора была дополнена как коллективным пользовани ем ресурсов и социальной сегментации внутри групп, так и военными действия ми и другими формами конфликтов между группами.
Результаты построений говорят о продуктивности подхода. Шкала времени, в рамках которой развивается история в простейших эволюционных стохасти ческих моделях, слишком медленна для того, чтобы быть совместимой с реаль ными траекториями человеческого развития, но введение локальных взаимодей ствий и коррелированности специфических действий в рамках конформистских тенденций сильно увеличит скорость изменений. Аналогично в отсутствие внут ригрупповых институтов, таких как коллективное пользование ресурсами и сег ментация, условия для развития индивидуальных поведенческих характеристик, выгодных для группы, достаточно убедительны. Когда же предполагается, что эти два практически важных аспекта взаимоотношений людей развиваются со вместно с индивидуальными характеристиками, это приводит к тому, что про цессы совместной эволюции все больше и больше принимают за возможное описание истории человечества.

Часть IV
заключение