

1 СЕМЕСТР. Экономика. Микроэкономика. Поведение, институты и эволюция Самуэль Боулз / Микроэкономика. Поведение, институты и эволюция_Самуэль Боулз_2010 -576с
.pdfГлава 13. Совместная эволюция институтов и предпочтений 429
ления. Определим за βG ≡ βg + βi влияние частоты появления характеристики в группе на среднее число реплик (разница между числом реплик, создаваемых
индивидом в группе, полностью состоящей из обладателей данной характери |
|
стики, и в группе без нее, равна βG). Значит, используя вышеупомянутые опреде |
|
ления, βi = -c, βg = b, βG = b - c. Следуя Прайсу (Price, 1970), |
|
πDp = var(pj)βG + E{var(pij)}βi |
(13.2) |
или |
|
πDp = var(pj)(b - c) - E {var (pij)}c,
где π — среднее по популяции число реплик (которое я нормирую единицей) и оператор ожидания E{} означает средневзвешенную сумму по группам (в роли весов выступает относительный размер группы). Первое слагаемое — влияние отбора на уровне группы (положительное), а второе отвечает за влияние отбора
на уровне индивидов (отрицательное) (простой вывод из обоих эффектов содер жится у Боулза (Bowles, 2001)). Если исключить вырожденные случаи, такие как
нулевая дисперсия, то из выражения следует, что частота появления характери стик внутри группы будет стационарной, когда дисперсии двух слагаемых равны (предполагая, что все β и дисперсии двух слагаемых стационарны). Поскольку второе слагаемое отрицательно, то частота появления характеристики внутри всех выживших групп станет со временем падать. Но поскольку βG положи тельно, то данная тенденция перевесит более высокий уровень приспособления группы с бо2льшей частотой появления А.
Теперь условие стационарности для p (уравнение (13.2)) показывает, что
Dp = 0, когда |
|
|
|
|
var( pj ) |
|
|
|
|||
c |
= |
|
|
|
, |
(13.3) |
|||||
|
|
|
|
|
|
|
|||||
b E {var( pij )} + var( pj ) |
|
|
|
||||||||
и |
|
|
c |
|
|
var ( pj ) |
|
|
|||
Dp > 0 при — |
< |
|
, |
||||||||
b |
E{var ( pij )} + var ( pj ) |
||||||||||
|
|
|
|
|
|
|
|||||
Dp < 0 при — |
c |
> |
|
var ( pj ) |
|
. |
|||||
b |
E {var ( pij )} + var ( pj ) |
|
|||||||||
|
|
|
|
|
|
|
|||||
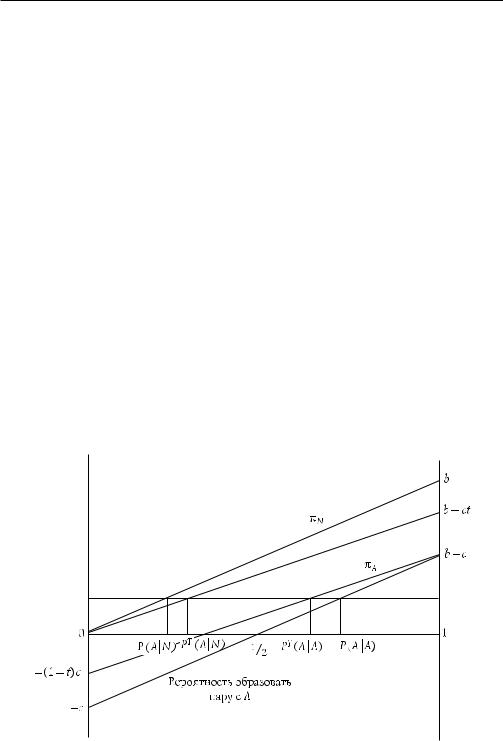
Левая часть — отношение выгод к затратам от альтруистических характе ристик. Правая часть — отношение межгрупповой дисперсии к сумме внутри групповой и межгрупповой дисперсии характеристики, что несложно показать (Bowles, 2001) и проиллюстрировать на последующем примере. Данное отноше ние измеряет разницу между вероятностью того, что альтруист образует пару с альтруистом P (A|A), и того, что он (неальтруист) образует пару с альтруистом P (A|N). Тогда
var ( pj ) = P(A A) −P(A N) = r. E{var ( pij )} + var ( pj )
Таким образом, отношение дисперсий — это мера степени сегментации по всей популяции, произошедшей не потому, что индивиды преднамеренно объ

430 Часть III. изменения: совместная эволюция институтов и предпочтений
единялись в пары внутри группы, а изза того, что популяция структурирована по группам. Равенство (13.3) показывает: для распространения альтруистиче ской черты в популяции необходимо, чтобы характеристика была тем дороже (относительно полученных выгод), чем больше дисперсия между группами (по отношению к дисперсии внутри группы). Из равенства (13.3) становится понят ной ключевая роль дисперсий между группами. Когда дисперсия между сред ними показателями группы равна нулю, вероятность того, что данный индивид встретит альтруиста, не зависит от типа самого индивида. Тогда групповой отбор не играет роли, и, таким образом, выгодная для группы характеристика станет распространяться, только если она не содержит издержек.
Соответственно, когда var (pij) = 0 j, все группы окажутся однородными
икаждый индивид образует пару только с индивидом своего типа независимо от состава всей популяции; внутригрупповой отбор отсутствует и влияние ока зывает только отбор межгрупповой. В таком (крайнем) случае можно сказать, что группа представляет собой цельную единицу отбора. Следовательно, сила
группового отбора будет зависеть от масштаба групповой выгоды относительно индивидуальных издержек (b и c в примере) и от того, насколько различается между группами средняя частота появления характеристики по отношению к внутригрупповой дисперсии характеристики.
Те, кто знаком с популяционной биологией, узнают в равенстве (13.3), за писанном в виде c/b = r, вариант правила Гамильтона для степени позитивного выбора подобных r, позволяющей характеристике «альтруизм» распространять ся даже в случае, когда эта характеристика редка. В данном отношении много уровневый отбор неотличим от эволюционного процесса, основанного на других формах выбора подобных (таких, как родственный отбор и иные формы внутри групповой сегментации, или кластеризации).
Поясним на примере. Популяция состоит из двух групп одинакового раз
мера, в которых доли альтруистов равны соответственно p1 = 3/4, p2 = 1/4, таким образом, общая доля равна p = 1/2. Из вышеприведенной платежной матрицы мы знаем, что выплаты альтруистов в каждой группе меньше выплат неальтруи стов. Значит, альтруисты в процессе репликации будут находиться в менее вы
годном положении. Это можно заметить, если посмотреть на функции выплат на рис. 13.2: для всех значений р выплаты неальтруистам превышают выплаты альтруистам (на время забудем про обозначенные пунктиром функции выплат). Но под влиянием группового отбора в группах с большой долей альтруистов вы
платы всем представителям группы станут выше, так что в таких группах аль труистическая характеристика может не исчезнуть. Чтобы найти значения b и c такие, чтобы p стала стационарной, необходимо приравнять средние показатели
приспособления для индивидов двух типов. Записываем pj для альтруистов из группы j (j {1, 2}), πij для приспособления игрока i из группы j (i {A, N}) и πi для среднего приспособления i. Используя равенство (13.1), приравнивая β0 = 0
изамечая, что p1 + p2 = 1, а группы имеют одинаковый размер, получаем, что для равенства уровней приспособления необходимо
πA = p1πA1 + p2πA2 = (1 - p1) πN1 + (1 - p2) πN2 = πN

Глава 13. Совместная эволюция институтов и предпочтений 431
Рис. 13.2. Эволюция альтруистической черты. Если структура популяции такова, что разность условных вероятностей образовать пару с A P(A|A) − P(A|N), как показано на рисунке, то p стационарна
или, подставляя уже имеющиеся данные,
πA = 34 (34 b −c )+ 41 (41 b −c )= 41 (34 b )+ 34 (4b )= πN .
Решая, получаем значения b и c, для которых Dp = 0, а именно c/b = 1/4. Если добавить требование, чтобы размер популяции оставался постоянным
(πA = 1 = πN), получаем b = 8/3, c = 2/3.
Аналогичным образом можно было просто применить равенство (13.2) и то,
что var(pij) = pj (1 - pj) = 3/16, j = 1, 2 и var(pi) = 1/16. Таким образом, используя равенство (13.2), получим
πΔp = b16−c − 163c ,
где при π ≠ 0 получаем c/b = 1/4 в качестве условия для Dp = 0, что совпадает с полученными ранее результатами. Далее воспроизводим равенство (13.3) и под ставляем эмпирические значения из примера, и, как и ожидалось, получаем
c |
= |
|
var( p j ) |
|
|
= |
|
1 |
16 |
|
= 1 |
4 |
. |
||||
b |
E {var( p |
ij |
)} + var ( p |
j |
) |
3 |
16 |
+ 1 |
16 |
||||||||
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Таким образом, для значений b >4c частота появления альтруистической ха рактеристики станет расти, превышая одну вторую в следующем периоде. Это происходит потому, что рост относительного размера группы, состоящей из большего числа альтруистов, перевешивает падение доли альтруистов в каждой группе. Распространение выгодных для группы, но убыточных для индивидов характеристик объясняется структурой популяции, поскольку альтруисты стре мятся объединяться в пары с другими альтруистами в среднем чаще, чем осталь

432 Часть III. изменения: совместная эволюция институтов и предпочтений
ные представители популяции (несмотря на случайный порядок объединения в пары внутри групп). Значит, вероятность того, что индивид встретит альтруиста, при условии, что он сам альтруист, равна
P(A|A) = (p1)2 + (p2)2 = 5/8,
в то время как вероятность для неальтруиста встретить альтруиста равна
P(A|N) = (1 - p1) p1 + (1 - p2) p2 = 3/8.
Разница между этими двумя условными вероятностями 1/4 — ожидаемое пре имущество, которое игроки получают от альтруистической характеристики пу тем более благоприятного перераспределения между группами. Можно пере писать равенства (13.3) в виде
c/b = P (A|A) - P (A|N) = 1/4.
Рисунок 13.2 показывает, как структура популяции может покрыть издерж
ки альтруистического поведения. В то время как выплата неальтруистам всегда превышает выплаты альтруистам при заданной вероятности встретиться с альтруистом, разница в условной вероятности встречи с альтруистом в зависи
мости от типа индивида (т. е. P (A|A) - P (A|N) = 1/4) покрывает этот недостаток. Пример показывает, как групповой отбор может способствовать распро странению характеристики, которая в ином случае не смогла бы сохраниться. Однако проведенный анализ не является полным. Уравнение Прайса дает пред ставление только о равновесии, но не обо всей динамической системе. Оно за дает условие стационарности p, но не объясняет изменение дисперсии, на кото ром основано изменение p. В большинстве биологических моделей механизмы, увеличивающие межгрупповую дисперсию (мутации, генетический дрейф), сла бые и в итоге попадают под влияние отбора наряду с миграциями между груп пами, делающими популяцию однородной. По этой причине считается слабым давление отбора на уровне группы среди животных. Тем не менее там, где эф фективный размер группы мал (например, среди сборщиков урожая) и где они часто делятся в ответ на увеличение своего размера либо напряженную внутрен нюю атмосферу, отклонения станут увеличивать дисперсию между группами. В любой, даже очень далекой от реальных условий эволюции человека модели единственным практическим способом определить, станет ли такое увеличение дисперсии достаточно большим, чтобы групповой отбор серьезно влиял на эво люцию, выступает построение модели, структурированной по группам популя
ции с разумными значениями параметров.
агентная модель многоуровневого отбора
В отсутствие двух введенных институтов на уровне групп — коллективного поль зования ресурсами и сегментации внутри группы — процесс отбора внутри группы строится (для группы j) с помощью стандартного уравнения реплика ционной динамики
Dpj = pj (1 - pj)(πAj - πNj) = pj (1 - pj)(-c). |
(13.4) |
Глава 13. Совместная эволюция институтов и предпочтений 433
Теперь представим, что внутри группы, как принято среди сборщиков уро жая и в других сообществах, установлено коллективное пользование ресурсами.
Индивид отдает некоторую долю добытого в «общий котел», — возможно, неко торые виды продуктов, как встречается среди аче (Kaplan & Hill, 1985), — сред
ства из которого затем делятся поровну между всеми представителями группы. Можно построить такой институт коллективного пользования, как линейная система налогообложения tj [0, 1), где налоги являются платежами, которые собираются с индивидов, а затем распределяется поровну между всеми членами популяции. Смысл состоит в том, чтобы сократить различие между выплатами
A и N, т. е. πAj - πNj = -(1 - tj)c. На рис. 13.3 изображены ожидаемые выплаты и влияние коллективного пользования ресурсами на разницу в выплатах индиви
дам двух типов в предположении, что все представители группы платят по одина ковой налоговой ставке t. Теперь разница в условных вероятностях встречи с А (в зависимости от типа индивида), равная ожидаемым выплатам, больше не рав на P (A|A) - P (A|N) = r*, как показано на рис. 13.2, а равна PT (A|A) - PT (A|N) = rT при rT < r*. Сравнивая два рисунка, можно увидеть, что r* = c/b, притом что rT = c (1 - t)/b. В результате если структура популяции была бы такой, как на рис. 13.3 (r*), и если присутствовал бы институт коллективного пользования ре сурсами (t > 0), то πA > πN и p возрастало бы.
Предположим, что вдобавок к институту коллективного пользования ресур сами группы сегментированы таким образом, что в процессе образования пар внутри групп A вероятнее всего станут взаимодействовать с A и N с N, чем при
Рис. 13.3. Коллективное пользование ресурсами ослабляет внутригрупповой отбор. Пунктирные линии функций приспособления отражают эффект института коллективного пользования ресурсами: альтруистическая характеристика может распространяться в менее строгих условиях, чем на рис. 13.2

434 Часть III. изменения: совместная эволюция институтов и предпочтений
случайном образовании пар. Предположим, что вероятность того, что A из груп пы j объединится с A, равна уже не pj, а sj + (1 - sj) pj > pj, а вероятность того, что N объединится с А, равна (1 - sj) pj < pj. Тогда (как и в гл. 7) определим sj ≥ 0 как степень сегментации в группе j, или разницу в условной вероятности встречи А с А и N с А при образовании пар внутри группы. Затем, абстрагируясь от налога (tj = 0): πAj - πNj = sjb - c. Сегментация снижает ожидаемую стоимость ущерба для альтруистов, потому что внутри заданной группы вероятность встречи с дру гими альтруистами распределена неравномерно, как и вероятность встречи N с представителем типа N. Если sj > c/b j, то в среднем А будут получать внутри каждой группы больше, чем N, и в итоге А будут распространяться как резуль тат внутри и межгруппового отбора. Значит, оба слагаемых в уравнении Прайса будут положительными. Чтобы сформулировать его в виде классической пробле мы группового отбора, мы предполагаем, что s < c/b, таким образом, А будут распространяться, если давление группового отбора будет достаточно сильным. Как и коллективное пользование ресурсами, сегментация достигается путем со глашения и происходит посредствам культурного отбора.
Учитывая сегментацию и коллективное пользование ресурсами, разница между ожидаемыми выплатами, получаемыми А и N, будет равна (1 - tj)(sjb - c). Таким образом, мы получаем
Dpj = pj (1 - pj)(1 - tj)(sjb - c), |
(13.5) |
из чего видно (сравнивая равенства (13.4) и (13.5)), что оба института замедляют групповой отбор, направленный против А. Это можно увидеть, заметив, что
∂ |
p j |
= −p j (1 − p j )(s jb −c), |
|
∂ |
∂t |
|
|
|
|
||
p j |
= p j (1 − p j )(1 −t j )b . |
(13.6) |
|
|
∂s |
||
|
|
|
Для pj (0, 1) оба выражения положительны. Это означает, что и сегментация, и коллективное пользование ресурсами оттесняют негативные последствия от бора против А. Заметим, что влияние каждого института больше, когда pj близ ко к одной второй и когда другой институт работает на низком уровне. Значит, в терминах выгоды от препятствия неблагоприятного для А отбора институты заменяют, а не дополняют: их выгодное влияние усиливается тем больше, чем меньше присутствие другого института.
Структура процесса корректировки описана на рис. 13.4 и в примечаниях к нему. Репликация индивидов вызвана мутациями, такими, что с некоторой малой вероятностью e потомок может стать А либо N с той же вероятностью. Институты, обозначенные за s и t, отличаются по группам, и они также имеют тенденцию развиваться. Когда возникает конфликт между группами, выигрыва ет группа с большими общими выплатами. Представители проигравшей группы умирают, а выигравшие занимают их места вместе со своими репликами. Новые представители подчиняются институтам той группы, от которой они произош ли. Институты могут также возникать вследствие стохастических изменений,

Глава 13. Совместная эволюция институтов и предпочтений 435
Рис. 13.4. Индивидуальные и групповые взаимодействия. Мы разделили n индивидуумов на g групп. При t = 0 все являются N. (1) Образование пар. В каждом периоде каждый
член группы случайно выбирает себе пару для игры с платежами, приведенными в тексте (в некоторых прогонах дополнено правилом коллективного пользования
ресурсами). При сегментации участник взаимодействует с себе подобным с вероятностью s, и выбор пары происходит случайно с вероятностью 1 – s. (2) Воспроизводство. Репли
ки текущего поколения образуют следующее поколение. Они производятся путем
копирования (с замещением) из состава текущей группы, причем вероятность быть скопированным равна доле члена в общем выигрыше группы. (3) Мутация.
Свероятностью e член следующего поколения не является репликой своего родителя,
аявляется A или N с равными вероятностями. (4) Миграция. С вероятностью m каждый
член следующего поколения присоединяется к случайно выбранной группе.
(5) Групповая конкуренция. С вероятностью k выбирается каждая группа, и среди выбранных происходит конкуренция в случайно составленных парах. Побеждает группа
с наибольшим общим платежом (очищенным от издержек коллективного пользования ресурсами и сегментации, если они есть). (6) Вторичное заселение и деление на группы.
Члены проигравшей группы заменяются репликами членов победившей группы, и в результате победившая группа (постоянно увеличиваясь) случайно разделяется на две новых группы (в симуляциях с коллективным использованием ресурсов или сегментации новые группы принимают институты победившей группы)

436 Часть III. изменения: совместная эволюция институтов и предпочтений
случайного увеличения или уменьшения t и s в каждом периоде. Как сегмен тация, так и коллективное пользование ресурсами приносят затраты группам, принимающим эти институты. У более сегментированных групп может не по лучиться извлечь выгоду из неоднородности групп или экономии от масштаба,
иколлективное пользование ресурсами способно ослабить стимулы к добыва нию ресурсов, которые впоследствии разделят между всеми. Ни одни из этих издержек формально не введены в модели, но, чтобы учесть их влияние, в модели
установлено, что средние выгоды для группы сокращаются на некоторую сумму, возрастающую и выпуклую по s и t.
ЮнгКьё Чой, Астрид Хопфензиц и я смоделировали искусственную попу ляцию, делящуюся на 20 групп. В табл. 13.2 указаны изначальные значения пара метров модели и диапазон их изменения, изученный нами. Ключевые параметры связаны с уровнем (случайной) миграции между группами, размером группы и частотой межгрупповых столкновений. Поскольку соревнования между груп пами устроены таким образом, что проигравший умирает, то мы решили, что группы вступают в соревнование (войну) один раз в течение жизни четырех по колений. Начальные значения были выбраны на основе наблюдений, о достовер ности которых я поговорю в предпоследнем разделе.
Мы моделировали каждую ситуацию без альтруистов и институтов в нулевой момент времени, чтобы посмотреть, распространятся ли они, если изначально
были редкими (процесс индивидуальных и институциональных мутаций при вносит изменчивость в популяцию). Базовый уровень приспособляемости (βo) равен 10, и потомки производятся пропорционально индивидуальной доле в
общей приспособляемости группы, таким образом в отсутствие сегментации и коллективного пользования ресурсами ожидаемое различие в выигрышах с = 1
иN производят на 10 процентов больше потомков, чем А.
|
|
|
|
Таблица 13.2 |
|
Ключевые параметры модели |
|
||
|
|
|
|
|
|
|
Изначальные значения |
|
Исследованный диапазон |
|
|
|
|
|
Среднее значение размера |
|
20 |
|
7 до 47 |
группы (n/g) |
|
|
|
|
Уровень миграции (m) |
|
0,2 |
|
0,1 до 0,3 |
|
|
|
|
|
Вероятность вступления |
|
0,25 |
|
0,18 до 0,4 |
в соревнование (k) |
|
|
|
|
Уровень изменений |
|
0,001 |
|
0,01 до 0,000001 |
(мутаций) (е) |
|
|
|
|
|
|
|
|
|
Примечание. Суммарный размер популяции равен n, и имеется g групп; m, k и e указаны для одного поколения. Другие параметры: выгода (b): 2; затраты (с): 1; базовые выплаты: 10. Мы изменяли размер группы, меняя n. По указанным в тексте причинам мы ограничили s так, чтобы оно не превышало 1/2, а t [0, 1]. Затраты, налагаемые этими институтами на группы, равны 1/2 (s2 + t2).
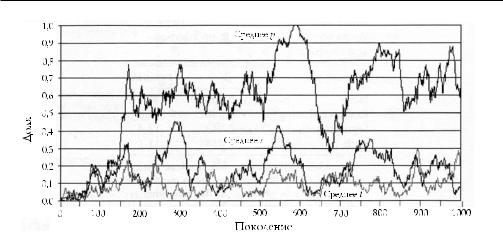
Построенная типичным образом популяция приведена на рис. 13.5. Ран ний рост p подкрепляется слу айным увеличением s и t (между периодами 100
Глава 13. Совместная эволюция институтов и предпочтений 437
Рис. 13.5. Динамическое взаимодействие между групповыми институтами и индивидуальным поведением. Рисунок отражает 1000 периодов с параметрами
из табл. 13.1. Средняя доля альтруистов — p, t и s отражает средний по 20 группам уровень коллективного пользования ресурсами и сегментации. Альтруизм и оба групповых института изначально редки. Рисунок четко отражает динамику, наблюдаемую на протяжении длительного периода во многих прогонах
и 150). Когда p достигает высоких уровней (в периоды с 532 по 588, например) s и t убывают, что естественно приводит к резкому снижению p. Последующий рост s и t происходит вследствие случайных событий.
Данный вид динамики наблюдается по следующей причине: когда популя ция разделена поровну на А и N, много групп разделены также примерно поров ну. В результате (из равенства (13.6)) выгода от замедленного внутригруппового отбора, получаемая при высоких уровнях t и s, в этой области максимальна. Тем не менее, когда p значительно выше 0,5, выгода от защиты А, достигнутая дей ствием институтов, меньше. Но институты обходятся дорого, так что, когда p велико, группы с достаточно высоким уровнем сегментации или коллективного пользования ресурсами могут проиграть в конфликтах с другими группами, а занятые ими места затем займут потомки победивших, которые испытывают на себе меньшие институциональные изменения. В результате, когда институты победивших работают в период жизни новых представителей популяции, s и t падают.
Чтобы изучить влияние институтов на процесс корректировки, мы оценили уравнение Прайса эконометрически, изучая влияние каждого института по от дельности (т. е. ограничивая к нулю только s, или только t, или ни один из параме тров). Используя данные для четырех симуляций по 10 тыс. поколений в каждой, мы построили регрессию, где наблюдаемая Dp была объясняемой переменной, а значения предыдущего периода для var (pj) и E {var (pij)} были объясняющими, где второй регрессор — средняя по всем группам внутригрупповая дисперсия. Коэффициенты при этих переменных — это βG и βi, оцененные из уравнения (13.2.). Как видно из табл. 13.3, последствием введения в комбинации коллектив ного пользования ресурсами и сегментации становится сокращение наполовину

438 Часть III. изменения: совместная эволюция институтов и предпочтений
последствий неблагоприятного для альтруистов внутригруппового отбора. Заме тим, что в отсутствие институтов оценка βi (0,102) очень близка к ожидаемому значению при условии, что базовый уровень приспособляемости равен 10 (та ким образом, N обладают преимуществом в приспособляемости в 10%). Оценка влияния межгруппового отбора βG (не показана) изменяется слабо в ответ на то, какие институты будут развиваться, и во всех случаях больше чем в четыре раза превышает влияние внутригруппового отбора. Средняя дисперсия внутри групп соответственно гораздо больше дисперсии между группами.
|
|
Таблица 13.3 |
Институты препятствуют неблагоприятному для альтруистов |
||
|
внутригрупповому отбору |
|
|
|
|
Институты |
βi |
t-статистика |
Ни одного |
-0,102 |
8,5 |
Коллективное пользование |
-0,080 |
16,6 |
ресурсами |
|
|
Сегментация |
-0,063 |
13,4 |
|
|
|
Оба |
-0,055 |
11,2 |
|
|
|
Примечание. В столбце βi указаны МНКоценки коэффициента при среднем по группам значении pj (1 - pj), используемого в качестве регрессора для Dpj (в качестве другого регрессора выступает дисперсия между группами, т. е. var (pj)). Последний столбец — абсолютное значение tстатистики.
Заметим, что мы можем переписать уравнение (13.2), т. е. условие того, что Dp = 0, в виде
|
β |
|
|
var ( pj ) |
|
|
||
− |
i |
= |
|
|
|
|
, |
|
β |
E {var ( p |
ij |
)} |
|||||
|
|
|
||||||
|
G |
|
|
|
|
|
||
где Dp > 0, если отношение дисперсий превышает отношение размера эффектов внутригруппового отбора и межгруппового отбора, и наоборот. Наблюдаем ли мы такие же результаты в построенной нами модели? Используя эконометри ческие оценки влияния внутригруппового и межгруппового отбора, указанные в табл. 13.3, так же, как и средние отношения дисперсий, наблюдаемые при нашем построении, мы получаем результаты табл. 13.4. Исключая действие институтов, мы видим, что отношение влияния внутригруппового отбора к межгрупповому, -βi /βG, почти в два раза больше отношения дисперсии между группами к дис персии внутри групп. Следовательно, если популяция находилась бы на уровне этих средних и полученных с помощью оценки значений, то Dp стало бы от рицательным. Значит, неудивительно, что в построенной модели, на которой и основаны эти оценки, среднее значение p равно 0,06. Тем не менее если действие обоих институтов не ограничено, то отношение дисперсий равно отношению влияний; это означает, что влияние внутригруппового неблагоприятного для А отбора в точности перевешивается влиянием межгруппового отбора, способ ствующего распространению типа А. В построенной модели, на которой основа ны эти оценки, среднее значение p равно 0,51.