

SELECTION_last_file
.pdf251
і селекційні аспекти використання цієї ознаки в програмах удосконалення ліній і порід тварин. Вони полягають в тому, що певний рівень статевого диморфізму в поєднанні з багатоплідністю і масою гнізда на час відлучення забезпечує більш повну реалізацію генетичного потенціалу продуктивності тварин, можливо, через вищі компенсаторні можливості росту і розвитку, характерні для особин, які походять із гнізд, ліній, родин із високим рівнем статевого диморфізму. Виходячи з цих передумов включено як ознаку добору рівень статевого диморфізму в лініях і родинах у програми селекції свиней на підвищення відгодівельних і м’ясних якостей.
Рівень статевого диморфізму в популяціях свиней обумовлений великоплідністю поросят і материнським впливом. Добір маток за рівнем статевого диморфізму гнізд пов’язаний з їх диференціацією за репродуктивними якостями. За однократного добору (у F0) найбільш високі показники багатоплідності, маси гнізда на час відлучення встановлені для маток класу М+ (відповідно 11,75 голови, 161,00 кг). Але в наступних генераціях без проведення добору спостерігається елімінація відмінностей і регресія ознак, у яких кращими за репродуктивними якостями були матки класу М-. Тому показник рівня статевого диморфізму слід оцінювати і вести добір ремонтного молодняку за ним в кожній генерації.
Співвідношення статей у гніздах, з яких вони походять.
Незаперечним є факт, що в окремих випадках у багатоплідних тварин співвідношення статей значно відхилюється від очікуваного 1:1. Значна кількість свиноматок й інших тварин за весь період використання дають в приплоді переважно (70% і більше) або виключно (100%) потомків однієї статі – самців чи самок. Одним із теоретичних підходів до пояснення цього явища може бути фертильність Х і У-сперміїв, обумовлена різною імунологічною сумісністю.
Існує думка (Манін В., Білоус Н., Вольницька Л., 1983), що різний антигенний склад Х і У сперміїв може бути підставою для селективного розмноження і регуляції співвідношення статей. Тому факти зміщення
252
співвідношення самців і самок від теоретично очікуваного мають місце і незважаючи на те, що спадково цей феномен не передається, заслуговують на увагу. У свинарстві є дані про різні показники відтворної здатності свинок, що походять з гнізд із різним статевим співвідношенням (Дмитриєва Г.Л., Филозопенко Л.І, Набродова Н.М., 1990).
Питання співвідношення статей у потомстві тварин і птиці давно цікавить дослідників щодо отримання переважно особин тієї статі, яка має більш високі продуктивні властивості. Особливого значення ця проблема набуває за необхідності отримання синів-плідників від видатних батьків із метою підвищення їхньої племінної цінності в стаді, популяції. Проблеми регуляції статі, в основному, можуть бути вирішені з використанням сучасних методів ембріогенетики, а селекційні аспекти в цьому випадку неефективні.
У дослідженнях впливу співвідношення статей на наступні репродуктивні якості свинок встановлено тенденцію до більш високих показників багатоплідності і молочності тих свиноматок, що походять із гнізд, у яких переважає чисельність самок.
Наукове розв’язання цієї проблеми дасть можливість прискорити селекційний прогрес за рахунок:
−отримання удвічі більшої кількості ремонтних плідників від високопродуктивних матерів, що дозволить підвищити селекційний диференціал їх добору за походженням;
−здійснення більш жорсткого добору жіночих особин до
племінного ядра завдяки його зменшенню в 1,8...2 рази у разі одержання тільки свинок.
Співвідношення статей у гніздах свиней має селекційно-технологічне значення. Матки, які походять із груп з співвідношенням у гніздах самок і самців 33%♀×67%♂, мали вищі, стійкі показники репродуктивних якостей (маса гнізда в 2-місячному віці на рівні 163,67 кг) порівняно з групою маток із співвідношенням 67%♀×33%♂ (147,89 кг). Кращі відгодівельні якості у
253
тварин двох суміжних генерацій спостерігались у потомків групи маток 33%♀×67%♂. Свинки мали менший вік досягнення живої маси 100 кг (231,6 доби у F1 і 221,45 доби у F2) та вищий рівень середньодобового приросту (522,54 і 516,51 г). Отримані результати свідчать про можливість підвищення відгодівельних якостей свиней шляхом добору у товарних господарствах підсвинків від маток із переважанням у гніздах частоти кнурців.
Вирівняність гнізд на час опоросу. У свинарстві досить ефективною важливою селекційною ознакою слід вважати критерій визначення вирівняності гнізд свиноматок за великоплідністю (Березовський М.Д., Ломако Д.В., 2001). Характеризувати цю ознаку слід через коефіцієнт мінливості живої маси поросят в гнізді під час народження (Сv), за лімітами (min-mах). Але найбільш точно вирівняність гнізд характеризує запропонований критерій оцінювання через індекс. Наочно розподіл поросят у вирівняних і не вирівняних гніздах за класом розподілу живої маси наведено на рис. 4.3.
Рис.4.3. Розподіл новонароджених поросят за живою масою у різних
гніздах
254
Вирівняність новонароджених поросят має високу позитивну кореляційну залежність із вирівняністю гнізда на час відлучення (r=0,75). Доцільність вибору показника вирівняності гнізда як селекційної ознаки підтверджується тим, що вона має високий рівень успадковуваності (h2=0,41) та достатню мінливість (Сv=39,44%), що створює умови для ефективного добору гнізд свиноматок. Перш за все, цю ознаку слід ввести до програми удосконалення материнських родинних форм.
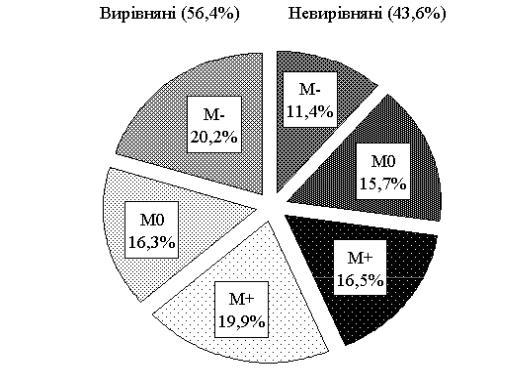
Однією з експериментально обґрунтованих ознак селекції свиней на підвищення репродуктивних і відгодівельних якостей є добір за індексом вирівняності гнізда. Маткам із вирівняних гнізд властива значно вища збереженість приплоду, жива маса гнізда на час відлучення поросят була на рівні 143,1…192,2 кг порівняно з 86,9…158,8 кг для невирівняних гнізд (Р<0,001). Вищі показники маси гнізда на час відлучення мали матки з співвідношенням 50%♀×50%♂: у вирівняних гніздах − 159,71 кг (Р<0,01) і
невирівняних − 153,08 кг з середньою масою одного поросяти 16,73 і 15,83 кг відповідно.
На підставі результатів використання моделі Т.Бріджеса встановлено, що новонародженим поросятам із невирівняних гнізд та з низькою живою масою, в основному, властиві більш високі показники кінетичної швидкості росту, але значно менші значення експоненційної швидкості у заключний період вирощування, тому вони суттєво поступаються за живою масою у 10-місячному віці і відгодівельними якостями тваринам з більшою індивідуальною великоплідністю (рис.4.4).
Чотирифакторним дисперсійним аналізом встановлено суттєвий вплив багатоплідності, вирівняності гнізда, статі та великоплідності на подальший ріст і відгодівельні якості поросят. Частка впливу багатоплідності на мінливість живої маси − на рівні 2,98% (у загальній дисперсії) і 14,66% − на показники середньодобового приросту. Вирівняність гнізд переважно впливала на вік досягнення живої маси 100 кг (11,73%). Максимальний вплив на рівень живої маси в 10-місячному віці має індивідуальна великоплідність

255
(25,50%). Спостерігається суттєва взаємодія між багатоплідністю маток та вирівняністю гнізда (10,17%).
Розробленими рівняннями множинної лінійної регресії, виходячи із показників репродуктивних якостей маток, встановлено переважний вплив на рівень живої маси на час відлучення багатоплідності маток (β-коефіцієнт – 0,584), вирівняності гнізда (β-коефіцієнт – 0,387) і молочності маток (β-коефіцієнт – 0,212). Виходячи з цих рівнянь можна з високою точністю прогнозувати масу гнізда на час відлучення.
Рис.4.4. Вік досягнення живої маси 100 кг залежно від співвідношення
статей у гнізді та його вирівняності за великоплідністю поросят
Стрес. Поряд з традиційно використовуваними ознаками добору останнім часом у свинарстві набувають поширення методи встановлення адаптивності генотипу до умов середовища. Це обумовлено тим, що тваринництво майбутнього буде базуватись на принципах адаптивності,
256
отриманні економічно виправданої продуктивності. До того ж останнім часом проблема стресу, адаптації і функціональних порушень увійшла до найбільш актуальних проблем сучасної біології і медицини. Це пов’язано з тим, що зростаюча індустріалізація, урбанізація, прискорення темпів життя й інші фактори, що супроводжують науково-технічну революцію, пов’язані з впливом на організм тварин різноманітних стресорів.
Для більш детального аналізу явищ стресу запропоновано класифікацію залежно від рівня організації організму тварин. Ця класифікація базується на тому, що неспецифічні (стресові) реакції характерні для всіх ступенів розвитку органічного світу, в чому і проявляється єдність принципів еволюції. При цьому слід зазначити, що проблема стресу в тваринництві вирішується головним чином через аналіз механізмів виникнення стресових реакцій, класифікацію стрес-факторів, усунення і профілактику шкідливих наслідків стресу. Але ці питання не пов’язують із періодами росту і розвитку організму тварин, проявом гетерозисного ефекту. Оцінювання реакції тварин на стрес-фактори може бути одним із критеріїв їх селекційного оцінювання. Вважають (Грікшас С.А., 1998), що сучасні методи ведення промислового свинарства не відповідають природним фізіологічним особливостям свиней, які склались у філогенезі.
Інтенсифікація відгодівлі, цілеспрямована селекція на підвищення м’ясності, безвигульне утримання великими групами, раннє відлучення молодняку, значні коливання складових мікроклімату та інші фактори створюють значний тиск на організм, який перевищує природні адаптаційні можливості тварин. У результаті дії цих факторів у свиней не тільки знижується продуктивність, але й виникає захворювання, яке отримало назву
стресового синдрому (PSS – Porkine stress sindrome), а також синдром PSE,
пов’язаний з прискореним розпадом глікогену в м’язах, різким підвищенням концентрації молочної кислоти та зменшеним рH – з 6,8...7,0 до 5,4...5,8. Вказане значно ускладнює одночасну селекцію на підвищення м’ясності туш за високої якості м’ясо-сальної продукції.
257
При цьому спостерігається послаблення конституції тварин, гострі серцеві захворювання з дегенерацією скелетних м'язів, нервовою збудністю, зниженням якості свинини і відтворювальної здатності.
Існує думка, що причиною виникнення стресового синдрому (РSS) є інтенсивна селекція свиней на м'ясність, вирощування яких в умовах промислового утримання призвело до зниження резистентності, підвищення чутливості тварин до стресів.
У селекційній роботі використовують такі основні методи оцінювання тварин на стресостійкість:
–галатановий тест;
–вивчення активності ферментів, імуногенетичний шок;
–ДНК-технології для виявлення рецесивного гена стрес-чутливості. Раніше найбільш широко використовувався галатановий тест (Webb A.,
1987). Серед протестованих за цим тестом та імунологічним шоком тварин виявлено (Бірдус Л.В., 1998) відповідно 22,5 та 17,5% стрес-чутливих свиней великої білої породи, 29,5 та 21,8% тварин породи ландрас, 19,0 та 15,2% – дюрок. Свині великої білої породи, ландрас і дюрок істотно відрізняються за частотою поширення стресового синдрому. Найбільшого поширення він набув серед свиней м'ясної продуктивності, зокрема породи ландрас, і менше серед універсальних, тобто великої білої. Найбільш нестійкими до впливу “генотип-середовище” є репродуктивні якості. Погіршення відтворювальної здатності найчастіше проявляється у вигляді підвищеного прохолосту і втрати молочності у маток, народження великої кількості нежиттєздатних і мертвих поросят, збільшення тривалості опоросу, появи хвороб органів травлення, інфекційних хвороб і передчасного вибуття із стада маток і кнурів.
Широкі можливості відкриваються для діагностики стрес-синдрому свиней у разі використання ДНК-технологій. Картування геномних локусів, що визначають господарсько-корисні ознаки тварин, пошук варіантів генів, які забезпечують їх бажаний прояв, є пріоритетними завданнями
258
сільськогосподарської генетики (Балацький В., Метлицька О., Биндюг А., 2000; Ревенко О., Войтенко С., Метлицька О., 2001).
Але проведення генетичного маркування щодо стрес-синдрому свиней до недавнього часу було можливим лише за поліморфізмом генів ферментів і груп крові, тісно зчеплених з галатановим локусом (Mucha A., Blicharski T., Fecert R., 1998;). Лише після виявлення мутації в ріанодинрецепторному гені (RYR1), що входить до галатанового локусу, стало можливим використання ДНКдіагностичного тесту, який зараз найбільш прийнятний і поширений у багатьох країнах із розвиненим свинарством.
Популяційно-генетичні дослідження показали, що в породах велика біла, миргородська RYR1-T-алеля не виявлено. Суттєва концентрація цього рецесивного алеля характерна тільки для полтавської м’ясної породи свиней і дещо менше для української м’ясної. Виявлена також асоціація гену, що обумовлює стрес-чутливість, із таким важливим комплексним показником як життєздатність поросят раннього віку.
Розглядаючи методичні аспекти оцінювання і добору свиней за стресостійкістю, слід вказати, що останнім часом розроблені більш ефективні заходи порівняно з галатановим тестом. Так, використовуючи принципи стабілізувального добору, запропонували (Коваленко В.П., Іванов В.О., Задирко В.І., 1988) вести оцінювання загальної стрес-чутливості за реакцією на стрес-відлучення.
Проведені дослідження показали, що нормоване відхилення зміни живої маси на 10 добу після відлучення становить від –1,5 до +1,5σ. Тобто, існує велика генетична можливість ефективного добору. При цьому особини, які менше реагують на стрес-відлучення (не знижують живу масу, залишаються на досягнутій величині або збільшують її), мають у наступному періоді онтогенезу значно вищі відгодівельні і м’ясні якості. Для них характерна більша енергія росту до 6-місячного віку. Отже, слід зазначити, що запропонований метод, завдяки його доступності, може широко використовуватись у практичній селекції.
259
Таким чином використання ДНК-технології і оцінювання реакції тварин на стрес-відлучення можуть використовуватися як ефективний захід диференціації генотипів за репродуктивними і відгодівельними якостями. Це створює передумови для організації вирощування свиней у рівночисельних угрупованнях, що сприятиме підвищенню рівня продуктивності тварин і загалом забезпечуватиме більш ефективне ведення галузі свинарства.
Наведені ознаки продуктивності не рівнозначні і їх величина обумовлена складною взаємодією генетичних і неспадкових факторів. У зв’язку з переведенням свинарства на промислову основу в умовах концентрації виробництва до важливих селекційних ознак, окрім продуктивності тварин, стали відносити і ті ознаки, які не формують саму продуктивність, але впливають на неї опосередковано. Це стосується селекції свиней на підвищення конституційної міцності, покращання адаптаційної здатності і стресостійкості до негативних впливів. Ці якості спадково обумовлені і, таким чином, успіх у селекції забезпечується добором.
У той же час, на показники конституції і адаптації особливо сильний вплив має зовнішнє середовище. Слід зазначити, що критерії селекції цих біологічних властивостей недостатньо розроблені і потребують подальшого вивчення.
4.4. Популяційно-генетичні параметри ознак селекції
Виділяють п'ять основних положень, що визначають ефективність селекції: генетичну обумовленість мінливості ознак; точність оцінювання спадкових якостей, тобто генотипу; добір; підбір; частоту зміни поколінь.
Генетична обумовленість мінливості ознак. Вона залежить від природи цих ознак, їх еволюційного розвитку і значення щодо забезпечення життєздатності тварини і вигляду загалом. З урахуванням цього всі ознаки свиней ділять на три групи: кількісні, якісні, порогові.
Кількісні ознаки характеризують безперервну зміну мінливості, тобто між особинами відсутні чіткі межі за розвитком, прояву тієї або іншої ознаки, тому є можливість їх групувати в різні класи без врахування генетичного
260
розщеплення. Одна з особливостей більшості кількісних ознак − їх велика залежність від умов середовища, а прояв їх у фенотипі зумовлено взаємодією генотипу з середовищем.
За рахунок того, що кількісні ознаки формуються в різних умовах порізному, для характеристики популяцій, стад, груп тварин використовують не частоти генів і генотипів, а статистичні показники, за допомогою методів варіаційної статистики.
У свиней до кількісних ознак відносять: багатоплідність, великоплідність, масу гнізда і масу одного поросяти на час відлучення, скоростиглість, м'ясність та ін.
Якісні ознаки свиней в основному визначають життєздатність і адаптаційну здатність (конституціональні якості і стресостійкість). До цієї групи також відносять: масть, групу крові, білкові поліморфні системи і пов'язані з ними спадкові аномалії. Особливість цих ознак − їх висока генетична обумовленість, невелика залежність від умов середовища і висока повторність у поколіннях. Для характеристики популяції за цими показниками використовують частоти генів і генотипів.
Прояв порогових ознак залежить від дії спадкових факторів і факторів середовища. Вони характеризуються дискретною мінливістю. Як правило, це альтернативні ознаки (вони або існують у тварини, або їх немає).
Наприклад, тварина хвора або здорова (для характеристики стійкості до якого-небудь захворювання), мертвонароджена або живонароджена, дає потомство або безплідна.
Ймовірно, що модель дії генів із пороговим ефектом така:
-усі фактори хвороби (генетичні і середовища) об'єднуються в одну безперервну випадкову величину, яку називають схильністю;
-схильність до хвороби має нормальний розподіл і проявляється як постійна безперервна ознака в цій популяції;
-відсутні загальні для споріднених (із схожим генотипом) тварин ефекти впливу середовища, які викликають захворювання.