

Osnovy_botaniki
.pdfОСНОВЫ БОТАНИКИ
Впервые корневые клубеньки были описаны в 1866 г. русским ботаником М.С. Ворониным.
На корнях бобовых растений обитают бактерии из рода Rhizobium. Они проникают в кору через корневые волоски, затем они занимают специальные паренхимные клетки и усиленно размножаются. Клетки с бактериями давят на другие ткани, образуя бактериоидную ткань. Эта ткань образует выросты на поверхности корня, которые называются корневыми клубеньками. Бактерии, которые вызывают появление этих клубеньков, называются клубеньковыми. У разных растений размеры и количество корневых клубеньков различно. Клубеньковые бактерии способны усваивать свободны азот воздуха, который обычно заполняет межклетники паренхимы корня и используется растениями.
Бобовые играют важную роль в обогащении почвы азотистыми веществами. Люпин, клевер используется в с/х в качестве удобрения (их запахивают). Эти растения называют сидератами.
Другие метаморфозы корней
Корнеклубни (или корневые шишки) образуются на придаточных или на боковых корнях (например, георгины, чистяк, ятрышник и др.), воздушные корни характерны для некоторых тропических растений, в частности, орхидей. На нижних частях стеблей кукурузы образуются придаточные корни, придающие растению устойчивость и играющие роль подпорки для всего растения. Корни-присоски (гаустории) развиваются у растений паразитов - повилика, омелы и др. Корневые отпрыски развиваются из придаточных почек корней у яблони, вишни, сливы, хрена, одуванчика и многих других растений. У мангровых, обитающих на береговых отмелях океанов в тропиках, развиваются ходульные корни, защищающие растения от затопления приливами.
8.6 Микориза
Слово «микориза» в переводе означает грибокорень от греческого «микес» – гриб, «ридза» – корень.
Грибы поселяются на корнях как древесных, так и травянистых растений. У луговых, лесных растений почти всех видов на кончиках корней образуется микориза. К каждому виду растения приспособлен определенный вид гриба. Многие микоризные грибы относятся к шляпочным грибам (белый, масленок, рыжик, подосиновик, подберезовик и т.д.).
Грибы используют у растений безазотистые органические соединения (углеводы растения-хозяина), которые образуются у зеленых растений за счет процесса фотосинтеза; сами грибы доставляют растению из почвы воду и минеральные вещества. Благодаря сильному ветвлению гиф гриба у корней, имеющих микоризу, сильно увеличивается поглощающая поверхность. Грибы так же передают растению частично органические вещества почвы, так же снабжают растения различными веществами-стимуляторами. Благодаря грибам высшие растения лучше развиваются и растут. Микоризные грибы к самостоятельной жизни не способны, а растения или развиваются плохо или
Полесский государственный университет |
71 |

ОСНОВЫ БОТАНИКИ
вовсе не растут (орхидеи). Это хорошо видно при акклиматизации некоторых растений, когда почва не содержит гриба-симбионта.
Существует три вида микоризы:
1)Эктотрофная;
2)Эндотрофная;
3)Эктоэндотрофная.
1)Эктотрофная микориза – гифы гриба оплетают кончики корней с наружной стороны и проникают в межклетники (у деревьев, кустарников). В случае эктотрофной микоризы кончики коротких корней оплетаются гифами
ввиде войлока, при этом главный корень не опутывается гифами гриба, и продолжает расти, углубляясь в почву. На кончиках корней с эктотрофной микоризой корневые волоски не развиваются, так как их функции выполняют гифы гриба, которые частично остаются снаружи и создают впечатление корневых волосков. Внешне корни с эктотрофной микоризой напоминают кораллы. На поперечных срезах корней хорошо видно, как гифы гриба через межклетники проникают до энтодермы. На кончиках этих корешков чехлик не образуются.
2)Эндотрофная микориза – гифы гриба развиваются внутри клеток коровой паренхимы (орхидные, вересковые, большинство травянистых растений). Анатомическое строение корешков с эндотрофной микоризой имеет свои особенности: наружный слой коровой паренхимы по межклетникам пронизаны гифами гриба, которые на поперечном разрезе напоминают сетку. Гифы эндотрофной микоризы развиваются и внутри паренхимных клеток, а иногда и внутри эпидермальных клеток. При эндотрофной микоризе корневые волоски развиваются нормально.
3)Эктоэндотрофная микориза – у растений с эктотрофной микоризой гифы гриба проникают в клетки коровой паренхимы и там растворяются и
усваиваются в процессе жизнедеятельности клетки. При эктоэндотрофной микоризе от густой сети гиф гриба отходят тонкие разветвления, проникающие внутрь некоторых паренхимных клеток.
Эктотрофные микоризные грибы, кроме функции корневых волосков, выполняют функции минерализации органических веществ в ризосфере. Эндотрофные микоризные грибы принимают участие в обмене веществ внутри клетки, выделяют различные ферменты (особенно важна их роль в синтезе сахаров, с помощью которых возрастает сосущая сила).
Растения, осуществляющие почвенное питание с помощью микоризных грибов называются микотрофными. Некоторые ученые считают взаимоотношения грибов и растений взаимным паразитизмом.
Полесский государственный университет |
72 |

ОСНОВЫ БОТАНИКИ
ТЕМА 9: ПОБЕГ
ПЛАН:
9.1Строение и типы побегов
9.2Микроскопическое строение стебля
9.3Строение стебля голосеменных растений
9.4Строение стебля двудольных травянистых растений
9.5Строение стебля однодольных растений
9.6Понятие о стеле. Стелярная теория
9.1 Строение и типы побегов
Вегетативный побег – осевой орган высшего растения. Основная функция – воздушное питание растения. Состоит из стебля и листьев. И стебель, и листья образуются из верхушечной меристемы одновременно, поэтому они и составляют единый орган – побег. Помимо вышеуказанных органов побег включает почки – зачатки новых побегов, за счет почек осуществляется ветвление и рост побега. Листьями выполняется главная функция побега – фотосинтез, стебель осуществляет функцию – проведения и связи между листьями и корнями, несет на себе тяжесть всех листьев и боковых побегов (механическая функция). Дополнительными функциями стебля являются – запас питательных веществ, вегетативное размножение и т.д.
Строение побега
Участок стебля, где к нему прикрепляется лист, называются узлом, участок побега между двумя соседними узлами называется междоузлием. Угол, образованный между листом и участком стебля называется листовой пазухой. Различают два типа побегов:
1.укороченные – междоузлия сближены, например, качан капусты, плодушки плодовых деревьев.
2.удлиненные – узлы раздвинуты.
Для побега характерно явление метамерии, т.е. многократная повторяемость сходных структур. Метамер типичного побега включает узел с листом, боковой почкой (расположенной над узлом, в пазухе листа) и междоузлие.
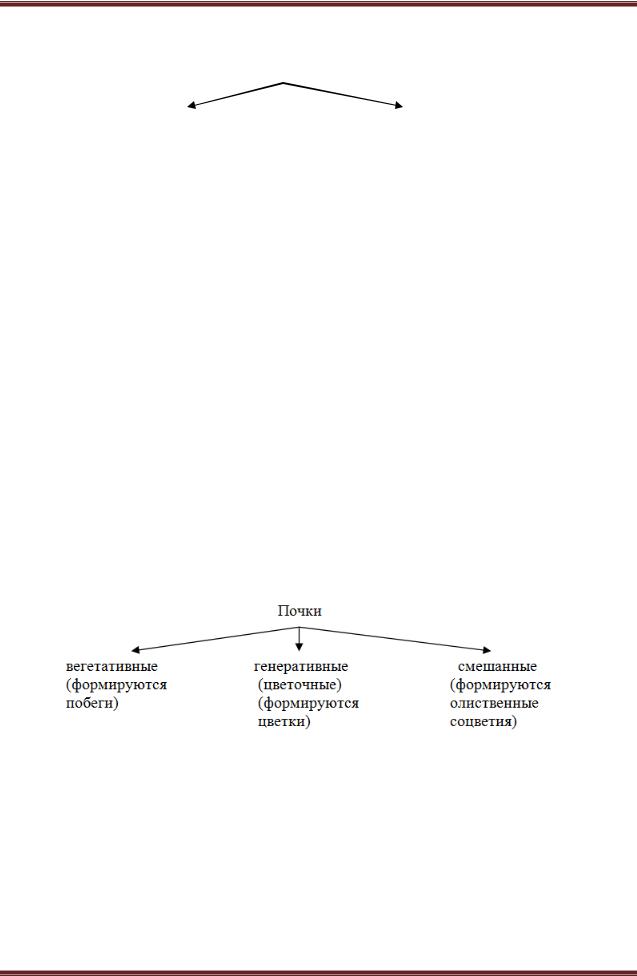
Почка
Почка – это зачаточный побег. Состоит из зачаточного стебля и зачаточных листьев, расположенных друг над другом. Нижние листовые зачатки прикрывают верхние листовые зачатки и конус нарастания. Это происходит за счет неравномерного роста.
Строение каждой почки максимально приспособлено к защите конуса нарастания. Особенно наружные листья, которые превращаются в почечные чешуи. Они могут быть пропитаны кутином, густыми клейкими или слизистыми веществами, смолами. Обычно чешуи пигментированы (защита от
Полесский государственный университет |
73 |
ОСНОВЫ БОТАНИКИ
перегрева), а также покрыты трихомами.
Почки (по расположению)
верхушечные (апикальные) |
боковые (пазушные) |
Большая часть боковых почек на следующий год после формирования распускаются и образуют новый побег – это активные почки.
Другие почки на следующий год не распускаются, но остаются живыми в течение многих лет. При утолщении стебля они нарастают, образуя в толще ствола скрытую ветку – это спящие почки. Они приходят в деятельное состояние при удалении вышерасположенной части стебля.
Почки возобновления – находятся на органах, не отмирающих в зимний период (т.е. на корневищах, у основания стебля). Зная особенности роста и развития почек, садоводы формируют крону плодовых и декоративных древесных растений, омолаживают старые деревья.
Придаточные почки – могут формироваться на всех частях растения (чаще на стеблях и корнях) – образуют корневые отпрыски, корневую поросль (у бриофиллиума – по краю зубчиков листа, образуя новые растеньица). В результате их деятельности легко размножаются листовыми черенками множество комнатных растений (бегония, герань, иризина, эхеверия). Придаточные почки берут начало от камбия, перицикла и других образовательных тканей.
У многих древесных растений от различных неблагоприятных факторов (от укуса насекомых, из-за повреждения тканей грибами, в результате действия химических веществ) из множества сближенных почек (придаточных) образуется «ведьмина метла».
Вегетативные и генеративные почки можно различить внешне, вегетативные имеют острую форму; генеративные и смешанные округлую форму и крупнее вегетативных.
Почки могут быть различной формы и размеров. Почки играют роль в приспособлении древесных растений к климатическим условиям. Почки могут располагаться: 1) одиночно; 2) сериально (одна над другой) – например, грецкий орех; 3) коллатерально (бокобочно) – например, чеснок, инжир. Само расположение почек бывает: очередное, или спиральное; супротивное.
Полесский государственный университет |
74 |

ОСНОВЫ БОТАНИКИ
Классификация жизненных форм растений по дат. уч. К. Раункиеру
(основана на положение почек)
•Фанерофиты – растения, почки возобновления которых, находятся высоко над поверхностью земли (деревья, кустарники);
•Хамефиты – растения, почки возобновления которых, расположены не высоко над поверхностью земли, зимой могут оказаться под снегом (полукустарники, кустарнички);
•Гемикриптофиты – растения, почки возобновления которых, находятся на уровне поверхности земли, надземная часть в неблагоприятный период отмирает (многолетние травы);
•Криптофиты - растения, почки возобновления которых, находятся ниже уровня поверхности земли, надземная часть отмирает (луковичные, клубневые, корневищные);
•Терофиты – однолетние растения, не имеющие почек возобновления, размножаются только семенами.
Листовой рубец
После опадения листьев на стебле остается след - листовой рубец, листовая подушка. На каждом листовом рубце хорошо видны небольшие точки – это следы идущих к листу проводящих пучков. Следы пучков на листовых рубцах называются листовыми следами. Листовые рубцы у различных растений различны и расположение их тоже различно. Листовые рубцы имеют форму треугольника, подковы, спирали, круга, полукруга; могут быть в виде сплошной линии, или состоять из отдельных точек расположенных различно. Форма и расположение листовых следов и листовых рубцов видоспецифична.
Фасциация
Фасциация – это явление, проявляющееся в уплощении или в сильном разрастании побега. Иногда встречается цилиндрическая фасциация. Причина фасциации – одновременное распускание нескольких сильно сближенных почек, которые в процессе роста образуют срастающиеся побеги. Фасциация наследуется и придает растениям декоративность. Появлению фасциации способствует обилие питательных веществ, частые дожди, повреждение верхушечной почки, гибридизация.
Ветвление
Из апикальной почки развивается главный побег (ось первого порядка), а из боковых почек – боковые побеги (ось второго порядка). На побегах второго порядка формируются боковые побеги третьего порядка, затем четвертого порядка. Стебель травянистых растений иногда не ветвится (у подсолнечника, кукурузы).
Благодаря ветвлению увеличивается общая масса надземных частей растений. У древесных растений в результате ветвления надземной части образуется крона. У деревьев главный стебель начинает ветвиться на определенной высоте от поверхности земли, и отрезок, не имеющий боковых разветвлений, называется стволом. У кустарников ветвление чаще всего
Полесский государственный университет |
75 |

ОСНОВЫ БОТАНИКИ
начинается от самой земли и ствол не выражен.
Эволюционно выработанные следующие способы ветвления: Дихотомической, или вильчатое – рост побега обеспечивается двумя
инициалиями верхушечной меристемы, растущими в одинаковом темпе и под углом в разные стороны. Это наиболее древний способ ветвления. Он характерен для водорослей, некоторых мхов, плаунов, папоротникообразных, некоторых голосеменных.
Моноподиальное ветвление – верхушечная почка образует главную ось, которая растет из года в год, главный стебель по сравнению с боковыми развит сильнее. Характерно для голосеменных растений, у покрытосеменных встречается редко.
Симподиальное ветвление – более поздний эволюционно тип ветвления. Его основу составляют дихотомический и моноподиальный типы ветвления.
При переходе дихотомического ветвления к симподиальному происходит сдвиг одного из апикальных инициалиев с конуса нарастания стебля в сторону другого инициалия. Активно растущий в длину инициалий заканчивает свой рост формированием такой же дихотомической развилки с неравноценным функционированием инициалиев.
При смене моноподиального способа – активизируется боковая почка, расположенная рядом с верхушечной почкой и подавляет своим ростом рост верхушечной почки, которая отмирает.
Внешне симподиальный побег напоминает моноподиальный, но состоит из симподиев (последовательного ряда осей). Симподиальное ветвление характерно для большинства покрытосеменных. При этом ветвлении рост растений в длину относительно небольшой, но формируется множество боковых ветвей, что способствует увеличению численности потомства – образуется много плодов и семян.
У большинства покрытосеменных моноподиальные и симподиальные типы ветвления комбинируются. Моноподиально ветвящиеся побеги обеспечивают рост, симподиальные – дают цветы и плоды – это комбинированное ветвление. У плодовых симподиальные побеги называются плодушками.
Ложнодихотомическое ветвление – особая форма симподиального ветвления. Апикальная почка отмирает или просто не развивается, рост продолжают две боковые почки, супративно расположенные под верхушечной почкой. Внешне напоминает дихотомическое ветвление. Характерно для гвоздичных, сирени, конского каштана.
Кущение – особая форма ветвления побегов (характерно для злаковых). У основания главного побега образуется группа боковых побегов. Ветвление происходит от укороченных узлов побега, расположенные под почвой или на уровне почвы, которые называются узлами кущения. Каждый побег в кусте на верхушке заканчивается соцветием.
Полесский государственный университет |
76 |

ОСНОВЫ БОТАНИКИ
По отношению к солнцу листья распределяются на побеге так, чтобы свет падал на все листовые пластинки.
При очередном (спиральном) расположении: листья на узлах размещены по одному. Если листья соединить по порядку образуется спираль. Прямая линия, соединяющая листья, расположенные друг над другом – называется ортостихой (от греч. «ортос» - прямой, «стихос» - ряд). Участки спирали между двумя листьями на одной ортостихе называется – листовым циклом. Число листьев в одном цикле и число витков в нем – видоспецифично.
Очередное листорасположение того или иного вида растения можно выразить дробью: в числители – число оборотов спирали, в знаменатели – число листьев. 1/2 - 1 оборот из 2 листьев (злаки), 1/3 – тюльпан, осока, 2/5 – широко распространено (груша, слива, яблоня).
Если сложить знаменатели и числители двух соседний дробей, то получим дробь следующего типа очередного листорасположения:
1/3 + 2/5 = 3/8, т.е. образуется математический ряд (1/2, 1/3, 2/5, 3/8). В
каждом листовом цикле угол, образованный между соседними листьями в
обороте – называется углом расхождения: 1/2 - < 1800, 1/3 - < 1200, 2/5 - <1440, 3/8 - < 1350.
При супротивном листорасположении на узлах друг против друга располагаются два листа (мята, крапива, сирень, жасмин и др.). Супротивно расположенные листья также образуют листовой цикл, то есть парно расположенные листья чередуются, образуя спираль.
При мутовчатом листорасположении на одном узле располагаются три и более листьев (вороний глаз, олеандр).
Главная роль листорасположения – это приспособление листьев к режиму освещения (приспосабливаясь, изменяется форма листьев, черешки, появляются их изгибы).
Расположение листьев, так чтобы они не перекрывали друг друга, для максимального использования лучей света – называется мозаикой листа, или листовой мозаикой.
Форма и типы побегов
Структура побега очень разнообразна и имеет ряд класификаций.
По форме поперечного среза: округлые (у большинства растений); трехгранные (осоки); четырехгранные (губоцветные); ребристые (зонтичные).
По наличию внутреннего содержимого: заполненные; полые
(соломина) – злаки, зонтичные.
По наличию листьев: облиственные; необлиственные (фиалка, лук, одуванчик, лилейные) – необлиственный цветонос – это последнее, единственно удлиненное междоузлие укороченного побега – стрелка.
Полесский государственный университет |
77 |

ОСНОВЫ БОТАНИКИ
Появление соцветия на стрелке называется – стрелкованием. Укороченная часть побега располагается у корневой шейки и образует прикорневую розетку (листья располагаются веерообразно).
По положению побега в пространстве:
•Прямостоячие – у большинства растений, хорошо развита механическая ткань (деревья, кустарники).
•Приподнимающиеся – у травянистых растений (яснотка, чабрец), механическая ткань развита слабее.
•Восходящие – растут под углом.
Выделяют 4 формы побегов с недостаточно развитой механической тканью:
•Вьющиеся и цепляющиеся побеги – сохраняют вертикальное положение при помощи опоры (лианы).
•Вьющиеся – против часовой стрелки (вьюнок полевой), по часовой стрелке (хмель),
•Цепляющиеся – имеют особые органы – прицепки, при помощи которых растение держится вертикально. Прицепки: усики (виноградная лоза), корни-прицепки (плющ).
•Лежачие, или стелющиеся – побеги не способны держаться вертикально (семейство тыквенных – огурцы, арбузы, дыни).
•Ползучие побеги – способны образовывать от узлов придаточные корни. Благодаря этому растения способны занимать большие территории (земляника, лапчатка).
9.2 Микроскопическое строение стебля
Стебель – надземная часть оси растения, один из его вегетативных органов. Стебель появился в процессе эволюции в связи с выходом растений на сушу. Полифункциональность стебля определила многообразие его внешнего облика и внутренней структуры. Степень развития различных тканей и их соотношение в стебле зависит от условий обитания и выполняемых функций, особенно доминирующих в данных условиях.
Отличительной чертой растений является их длительный рост за счет апикальных и латеральных меристем. В зависимости от того, за счет каких меристем происходит формирование стебля, различают первичное и вторичное его строение. Первичное строение характерно для стебля однодольных, вторичное – для голосеменных и двудольных.
Апекс стебля
Самая верхушка стебля называется конусом нарастания. Конус нарастания состоит из верхушечной, или апикальной, меристемы, ниже которой появляются небольшие бугорки – примордии (зачатки листьев). На уровне первых примордиев происходит формирование первичной структуры стебля. Строение апикальной меристемы неодинаково у разных групп растений, что привело к появлению различных точек зрения на строение апекса.
Полесский государственный университет |
78 |

ОСНОВЫ БОТАНИКИ
Теория апикальной клетки предполагает наличие одной клетки как функциональной единицы апикальной меристемы, которая управляет всеми процессами роста.
Теория гистогенов (Ганштейн, 1868). Согласно этой теории, тело растения возникает из массы меристематических клеток, которые расположены слоями (гистогенами). Самый наружный слой – дерматоген – является примордиальным эпидермисом, второй слой – периблема – дает начало коре, а третий – плерома – занимает внутреннюю часть оси.
Теория туники и корпуса (А.Шмидт, 1924). Согласно этой теории, в апикальной меристеме есть две зоны тканей – туника (слои периферических клеток) и корпус (масса клеток, покрытая туникой). Клетки туники делятся антиклинально, обусловливая рост верхушечной меристемы в поверхность. Клетки корпуса делятся во всех направлениях, обеспечивая объемный рост. Каждый слой туники и корпуса имеет свои инициальные клетки, то есть число слоев инициалей равно числу слоев туники и корпуса. Однако у растений трудно выделить четкую границу этих зон, поэтому предложено именовать тунику мантией, а группу клеток под ней – ядром. Эта концепция не приемлима для голосеменных, так как у них в точке роста просматривается зональность, основанная не только на плоских делениях, но и на цитологических и гистологических различиях.
К. Эсау (1969) выделяет в апексе дистальную зону, аксиальную, включающую ось, и еще две зоны, происходящие от аксиальной: первая – проксимальная, или внутренняя зона, расположенная непосредственно под дистальной, занимает центральное положение и превращается в сердцевину; вторая – периферическая, или наружная (фланговая меристема), окружает остальные зоны.
Изученные теории апекса говорят о большом разнообразии его строения у разных групп растений.
Развитие постоянных тканей стебля
Зона роста стебля более длинная, чем эта зона корня. Все производные апикальной меристемы претерпевают растяжение, после чего они специализируются. Меристематическая деятельность в периферических зонах и сердцевине затухает, но в конце клеток между ними она остается активной – это прокамбий, клетки которого ориентированы продольно. К прокамбию прилегают клетки перицикла. Его клетки могут оставаться паренхимными, но часто они удлиняются и становятся прозенхимными. Иногда их стенки утолщаются и одревесневают – превращаются в волокна.
Клетки прокамбия дифференцируются в элементы первичной флоэмы кнаружи и первичной ксилемы внутрь. Самые первые элементы называются протофлоэмой и протоксилемой, затем за ними дифференцируются элементы метафлоэмы и метаксилемы, которые как бы раздвигают первичные элементы. Таким образом, флоэма развивается в центростремительном направлении, а ксилема – в центробежном. Проводящие элементы протоксилемы – это трахеиды и длинные членники сосудов, оболочки которых имеют кольчатое
Полесский государственный университет |
79 |

ОСНОВЫ БОТАНИКИ
или спиральное утолщение, что облегчает растяжение этих элементов, так как одновременно с их формированием происходит и рост стебля. Проводящие элементы метаксилемы представлены лестничными и пористыми сосудами.
Участок паренхимы, остающийся в центре стебля. Называется сердцевиной, а полоса паренхимы к периферии от проводящего цилиндра – первичной корой. Самый внутренний слой первичной коры – эндодерма, но в стебле она выражена не четко. Еѐ клетки богаты крахмалом, который не расходуется растением – эти клетки называют крахмалоносным влагалищем, а крахмал – оберегаемым.
Если прокамбий полностью дифференцируются в элементы первичных проводящих тканей – стебель будет иметь первичное строение, что характерно для однодольных растений. У двудольных и голосеменных часть прокамбия превращается во вторичную образовательную ткань – камбий, который формирует вторичные постоянные ткани – стебель приобретает вторичную структуру.
В зависимости от того, как располагаются проводящие ткани, как в точке роста закладывается прокамбий различают стебли пучковые (у однодольных и травянистых двудольных) и непучковые (голосеменные и древесные покрытосеменные). Непучковые стебли могут формироваться или на основе прокамбиальных пучков или на основе прокамбиального кольца.
Строение непучкового стебля
( на примере древесного растения)
Развитие непучкового стебля происходит следующим образом. В апексе, в меристематическом кольце дифференцируются прокамбий, клетки которого образуют или тяжи (пучки), или кольцо. Прокамбий разграничивает основную паренхиму стебля на две зоны: центральную (сердцевину) и периферическую (первичную кору). Из клеток прокамбия образуются сначала протоксилема и протофлоэма, затем элементы метаксилемы и метафлоэмы, и в зависимости от расположения прокамбия первичные проводящие ткани будут расположены или в виде проводящих пучков, или сплошными кольцами по обе стороны от прокамбия. На этой стадии стебель древесного растения имеет первичную структуру. Часть клеток прокамбия превращается в камбий. Если прокамбий был в стебле в виде пучков, то из меристематических клеток между проводящими пучками сформируются камбиальные клетки (межпучковый камбий), которые, наряду с камбием пучков, образуют сплошное камбиальное кольцо. С этого момента стебель начинает формировать вторичную структуру. В результате деления камбиальных клеток первичные элементы будут сдвигаться к периферии.
В конце первого вегетационного сезона на поперечном срезе топография тканей будет следующая. В центре расположена сердцевина, которую окружает или сплошное кольцо первичной ксилемы, или отдельные еѐ группы. За первичной ксилемой расположена широким кольцом вторичная ксилема. Затем камбиальная зона, которая отграничивает древесину от луба. Проводящие ткани охватывают кольцо первичных механических элементов.
Полесский государственный университет |
80 |