

Osnovy_botaniki
.pdfОСНОВЫ БОТАНИКИ
Липидные капли, или гранулы, содержат различные смеси жиров. Образуются в гиалоплазме, а также в олеопластах. Размеры капель от 50 до 100 нм. Вначале липидные капли имеют водное содержимое, которое постепенно вытесняется отложением жиров. Затем гранулы окружаются мембраной (одиночным слоем липидов). Липидные капли называют сферосомами. Жирные масла в виде липидных капель откладываются в запас практически во всех клетках и являются основной формой запасных веществ большинства растений.
Кристаллы – представляют собой продукт метаболизма клетки, состоящий из солей многих органических и неорганических кислот. Они имеют специфическое строение и часто служат систематическим признаком. Кристаллы, содержащиеся в растениях, представлены в основном солями щавелевой кислоты, или оксалатами кальция, которые локализуются в вакуолях. У растений встречаются несколько видов кристаллов оксалата кальция: одиночные простые кристаллы ромбоэдрической, призмаэдрической, октаэдрической или удлиненной формы (сухие наружные чешуи лука, чеснока), стилоиды – колоновидные кристаллы (листья ириса), призмы – прямоугольные или пирамидальные (листья бегонии), рафиды – кристаллы в виде пучков игл (листья винограда, стебли традесканции), друзы – шаровидные образования, состоящие из многих мелких, сросшихся кристаллов (листья липы, плоды шиповника, злаковых, кора многих деревьев), кристаллический песок – очень мелкие кристаллы (паренхимные клетки бузины, пасленовых). Благодаря образованию кристаллов оксалата кальция происходит нейтрализация щавелевой кислоты (ядовитой).
При сильном разрастании кристаллы превращаются в идиобласты – одиночные клетки, включенные в какую-либо ткань, отличающиеся от клеток окружающей ткани формой, размерами, функциями. У некоторых растений образуется углекислый кальций (карбонат кальция), который пропитывает выросты клеточной оболочки, вдающиеся в полость клетки, образовывая гроздевидные образования – цистолиты (крапивные, тутовые). Также кристаллические включения могут состоять из кремнезема. Кристаллы играют важную роль в поддержании осмотического давления и кислотно-щелочного равновесия в клетке.
4.2 Теория симбиогенеза
В 1920-е, до Маргулиса теорию эндосимбиотического происхождения хлоропластов выдвинул русский ботаник Козо-Полянский (1924). Пятьдесят лет назад, в 1967 году, Линн Маргулис (Lynn Margulis) опубликовала развернутое изложение симбиогенетической теории, согласно которой эукариоты (организмы с клеточными ядрами) возникли в результате серии объединений разных клеток между собой. Современная поправка к этой теории гласит, что в основе становления эукариот, по-видимому, была не общая тенденция, охватившая многие эволюционные ветви (как предполагала Маргулис), а уникальное событие, приведшее к слиянию клеток археи и
Полесский государственный университет |
31 |

ОСНОВЫ БОТАНИКИ
протеобактерии. В результате образовалась сложная клетка с митохондриями, которая и стала первым эукариотом. Дальнейшие симбиогенетические события — например, захват водорослей, ставших хлоропластами, — действительно происходили много раз, но с возникновением эукариот как таковых они не связаны.
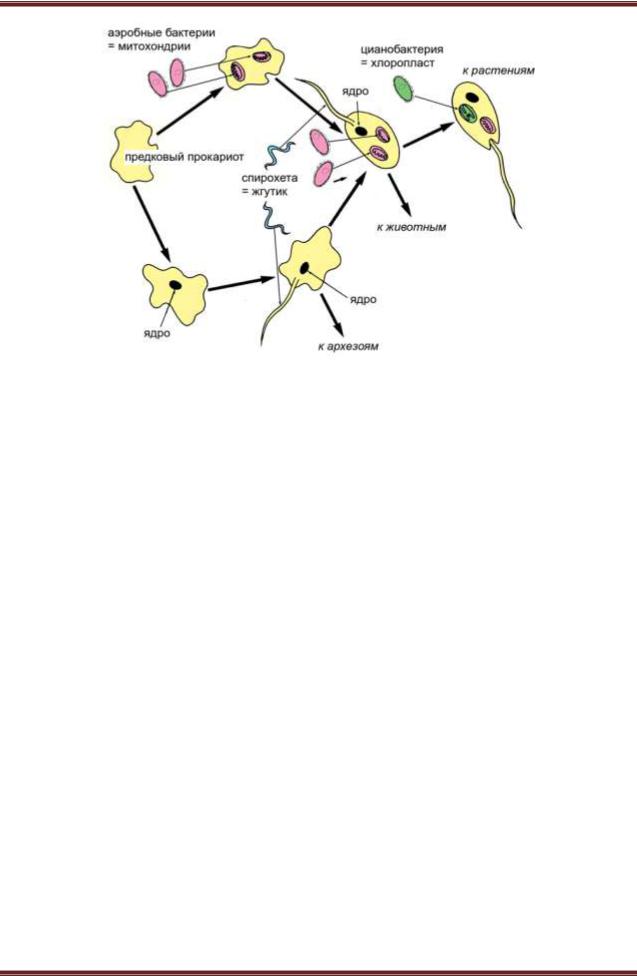
Выход в 1967 году статьи Линн Маргулис стал началом обновления биологических представлений, которое многие авторы расценили как смену парадигм — то есть, иными словами, как самую настоящую научную революцию. Суть интриги тут проста. Со времен Чарльза Дарвина биологи были убеждены, что основным способом эволюции является дивергенция — расхождение ветвей. Линн Маргулис была первой, кто сумел по-настоящему убедительно объяснить научному сообществу, что механизм некоторых крупных эволюционных событий, скорее всего, был принципиально другим. В центре интересов Маргулис оказалась проблема происхождения эукариот — организмов, клетка которых обладает сложной внутренней структурой с ядром. К эукариотам относятся животные, растения, грибы и многие одноклеточные — амебы, жгутиконосцы, инфузории и прочие. Маргулис показала, что ранняя эволюция эукариот вовсе не сводилась к дивергенции — она включала в себя слияние эволюционных ветвей, причем неоднократное. Дело в том, что по меньшей мере два типа эукариотных органелл — митохондрии, благодаря которым мы можем дышать кислородом, и хлоропласты, осуществляющие фотосинтез, — происходят не от того предка, что основная часть эукариотной клетки, рисунок 2. И митохондрии, и хлоропласты — это бывшие бактерии, изначально совсем не родственные эукариотам (протеобактерии в случае митохондрий и цианобактерии — в случае хлоропластов). Эти бактерии были поглощены клеткой древнего эукариота (либо предка эукариот) и продолжили жить внутри нее, сохраняя до поры собственный генетический аппарат.
Таким образом, эукариотная клетка — это, по выражению Маргулис, мультигеномная система. И возникла она в результате симбиоза, то есть взаимовыгодного сожительства разных организмов (точнее, эндосимбиоза, один из участников которого живет внутри другого). Соответствующие эволюционные ветви при этом, разумеется, слились. Такой взгляд на эволюцию получил название теории симбиогенеза.
Полесский государственный университет |
32 |
ОСНОВЫ БОТАНИКИ
Рисунок 2. – Пути эволюции древнейших эукариот, согласно взглядам Линн Маргулис
Примечание: Здесь показаны три симбиогенетических события — приобретение митохондрий, жгутиков и хлоропластов, — которые могли происходить в разных эволюционных ветвях в разном порядке. Тем не менее «верхний» и «нижний» пути эволюции ведут к одному и тому же состоянию: клетке, имеющей и ядро (происхождение которого тут оставлено «за кадром»), и митохондрии, и жгутики. У предков растений к этому добавляются еще и хлоропласты. Прокариотами называются все клеточные организмы, кроме эукариот, архезоями — гипотетические эукариоты, никогда не имевшие митохондрий.
Доказательства:
-разница в строении внутренней и наружной мембраны митохондрий -наличие в митохондриях собственной кольцевой ДНК (как у бактерий),
которая содержит гены для определенных митохондриальных белков -наличие в мембране собственного белок-синтезирующего аппарата,
причем рибосомы в нем прокариотного типа -деление митохондрий происходит простым бинарным путем, либо
почкованием и не зависит от деления клетки.
-Несмотря на определенную независимость митохондрии, находятся под контролем эукариотной клетки. Например, в гиалоплазме синтезируется некоторые белки, необходимые для нормального функционирования митохродний, и некоторых белковых факторов, которые регулируют деление митохондрий.
-ДНК митохондрий и пластид, в отличие от ДНК большинства прокариот, содержат интроны.
В собственной ДНК митохондрий и хлоропластов закодирована только часть их белков, а остальные закодированы в ДНК ядра клетки. В ходе эволюции происходило «перетекание» части генетического материала из
Полесский государственный университет |
33 |

ОСНОВЫ БОТАНИКИ
генома митохондрий и хлоропластов в ядерный геном. Этим объясняется тот факт, что ни хлоропласты, ни митохондрии не могут более существовать (размножаться) независимо.
Не решѐн вопрос о происхождении ядерно-цитоплазматического компонента (ЯЦК), захватившего прото-митохондрии. Ни бактерии, ни археи не способны к фагоцитозу, питаясь исключительно осмотрофно. Молекулярно-биологические и биохимические исследования указывают на химерную архейно-бактериальную сущность ЯЦК. Как произошло слияние организмов из двух доменов, также не ясно.
Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 году Андреас Шимпер, показавший их саморепликацию внутри клетки. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришѐл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников.
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии
Жгутики и реснички
Линн Маргулис предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путѐм деления, а путѐм достраивания нового органоида рядом со старым.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами.
Полесский государственный университет |
34 |

ОСНОВЫ БОТАНИКИ
ТЕМА 5: ТКАНИ РАСТЕНИЙ. ОБРАЗОВАТЕЛЬНЫЕ И ОСНОВНЫЕ ТКАНИ
ПЛАН:
5.1Понятие о тканях и их классификация
5.2Меристематические (образовательные) ткани
5.3Основные (паренхимные)ткани
5.1 Понятие о тканях и их классификация
В процессе эволюции дифференцировка тела растений сопровождалась дифференцировкой и усложнением внутренней структуры. Возникли группы клеток, которые выполняют определенную функцию. Еще в 1671 г. Н. Грю предложил назвать их тканями. Он, а также М. Мальпиги дали понятие об основных морфологических особенностях разных групп клеток. Те клетки, которые разрастаются более или менее равномерно по всем направлениям и диаметр которых примерно одинаков или соотношение длины и ширины не превышает 2 – получили название паренхимных. Такие клетки имеют изодиаметрическую или таблитчатую форму. Клетки, у которых длина более чем в 2 раза превышает ширину, были названы прозенхимными. Обычно они входят в состав проводящей и механической ткани. Это значит, что с морфологической точки зрения растения состоят всего из 2 типов клеток. Но в дифференцированном теле высших растений возникли группы клеток, которые похожи не только по форме, но и по функциям. Такие устойчивые комплексы клеток, которые похожи по происхождению, строению, выполняют определенную функцию и занимают определенное место в теле растений, называются тканями. Существуют разные типы тканей и разные подходы к их классификации:
1.По форме клеток, которые образуют ткань – ткани паренхимные
(основная паренхима, эпидермис) и прозенхимные (проводящие элементы, механические волокна).
2.По физиологическому состоянию клеток – ткани мертвые и живые.
Вмертвых отмирает цитоплазма, но клеточная оболочка сохраняется и продолжает играть важную роль (трахеиды, сосуды, древесинные волокна). В клетках живых тканей есть цитоплазма, и наблюдается циклоз.
3.По степени дифференциации клеток — ткани недифференцированные (меристематические) с высокой тотипотентностью — способностью к реализации всей генетической программы организма, и ткани постоянные, дифференцированные по функциям (покровные, основные, проводящие).
4.По времени и особенностям образования — первичные, вторичные и третичные. Первичные образуются из первичных, зародышевых или апикальных меристем (эпидермис, колленхима). Вторичные – из вторичных меристем или из постоянных тканей (вторичные проводящие ткани, камбий,
Полесский государственный университет |
35 |

ОСНОВЫ БОТАНИКИ
феллоген). Третичные образуются благодаря формированию перидерм и отмиранию всех расположенных между ними тканей.
5.По степени сложности выделяют ткани простые, состоящие из однотипных клеток (колленхима, паренхима) и сложные, образованные из клеток нескольких типов (ксилема, флоэма, перидерма).
В определенной степени имеют смысл все эти подходы к классификации. Однако наиболее рациональной и признанной является классификация тканей на основе анатомо-физиологических принципов, т. е. на основании строения тканей и выполняемых ими функции. С учетом этих признаков у растений выделяют 6 типов тканей:
1.Образовательные, или меристемы.
2.Покровные, или пограничные.
3.Механические, или арматурные.
4.Проводящие.
5.Основные (паренхимные).
6.Выделительные структуры (сборная группа).
5.2 Меристематические (образовательные) ткани
Тело растений формируется в результате деятельности образовательных тканей, или меристем (от греч. meristos – делимый). Клетки меристем способны к активному митотическому делению в течение всей жизни растения или достаточно длительное время, чем отличаются от образовательных тканей животных. Меристемы сохраняются столь длительный срок благодаря тому, что имеют меристематические клетки, образующие тело растений. Эти клетки имеют вид многогранников (обычно граней 14). Они производят подобные себе клетки основной меристемы, делятся неограниченное количество раз, как правило, параллельно граням, в разных плоскостях. Остальные, дочерние клетки, делятся ограниченное число раз и, в конце концов, пре вращаются в постоянные ткани – проводящие, покровные, механические,основные и т. д.
Классификация меристем основана на их положении в теле растения и на происхождении. По местоположению различают верхушечные, боковые и вставочные меристемы, по происхождению – первичные и вторичные.
Верхушечные (апикальные) меристемы располагаются на верхушке побегов и на кончике всех молодых корешков. Они обеспечивают рост побегов и корней в длину (высоту), их ветвление. Верхняя часть апикальной меристемы представлена инициалиями (единственной клеткой – у хвощей, многих папоротников; многоклеточной структурой – у семенных растений). Инициалии и ближайшие к ней клетки относят к зоне промеристемы. Вслед за ней идут ткани уже частично дифференцированные: в стебле – туника (образующая эпидермис и часть первичной коры) и корпус (образующий центральный цилиндр); в корне – дерматоген (образующий ризодерму), периблема (образующая первичную кору), плерома (образующая центральный цилиндр). При ветвлении каждый боковой побег и каждый боковой корень также обязательно имеют верхушечные меристемы со своими инициалиями.
Полесский государственный университет |
36 |

ОСНОВЫ БОТАНИКИ
Боковые (латеральные) меристемы располагаются параллельно боковой поверхности органа. В стеблях, корнях латеральные меристемы располагаются цилиндрическими слоями, имеющими вид колец на поперечных срезах. Боковые меристемы делятся на первичные (возникают непосредственно под апексом и тесно связаны с апикальной меристемой) – прокамбий и перицикл, и вторичные, которые образуются из первичных, либо из клеток постоянной ткани путем их дедифференциации (т.е. обратного преобразования в меристему) – камбий и пробковый камбий (феллоген). Боковые меристемы обеспечивают рост органов в толщину. Часто (у большинства однодольных растений) вторичные меристемы отсутствуют, и тогда все тело растения образовано одними лишь первичными меристемами.
Вставочные (интеркалярные) меристемы расположены у основания междоузлий. Они относятся к верхушечным меристемам, следовательно, по происхождению являются первичными, клетки которых надолго задержались в эмбриональном состоянии среди специализированных тканей, поэтому их ещѐ называют остаточными. От остальных меристем вставочные отличаются тем, что в них имеются некоторые дифференцированные элементы (например, проводящие) и нет инициальных клеток. Деятельность этих меристем обусловливает рост органов в длину после прекращения верхушечного роста. Участки интеркалярной меристемы имеются в черешках, пластинках листьев, цветоносах.
Раневые (травматические) меристемы возникают в любой части растения в результате повреждений. Клетки, окружающие пораненный участок, в результате дедифференциации приобретают способность к делению
иобразуют особую раневую ткань – каллюс, которая постепенно переходит в раневую пробку.
Согласно генетической классификации в зависимости от происхождения
ивремени появления в онтогенезе различают две группы меристем: первичные и вторичные. Первичные меристемы берут начало с момента деления зиготы и образования зародыша. До тех пор, пока зародыш не станет довольно крупным шаровидным телом, делятся все клетки и в разных направлениях. С момента образования боковых элементов зародыша – семядолей – происходит первая локализация меристем на двух полюсах. Между семядолями дифференцируется конус нарастания стебля, а на противоположной стороне – конус нарастания корня, т. е. формируются верхушечные, или апикальные (от лат. apex – вершина) меристемы. Именно за счет их деятельности корень и побег нарастают в длину. При ветвлении каждый боковой корень и каждый боковой побег имеют свои апикальные меристемы и инициали, что позволяет им, расти в длину. Апикальные меристемы обусловливают так называемый первичный рост в длину.
Кпервичным меристемам относятся также вставочные, или интеркалярные меристемы. Они отделяются от верхушечной меристемы, между ними развиваются зоны постоянных, дифференцированных тканей. В интеркалярных меристемах отсутствуют инициальные клетки, они со
Полесский государственный университет |
37 |

ОСНОВЫ БОТАНИКИ
временем прекращают делиться и превращаются в дифференцированные ткани. Интеркалярные меристемы находятся в узлах побега (особенно долго у злаков), в основании тычинок, листовых зачатков, черешков, цветоносов. Вставочные меристемы характерны для тех растений, у которых часто повреждаются апексы, или на верхушке закладываются зачатки цветков или соцветий.
На конусе нарастания очень рано дифференцируются производные апикальной меристемы – протодерма или дерматоген, прокамбий и основная меристема. Еще в ходе эмбриогенеза они формируют три первичные системы тканей: покровную (или дерматогена), проводящую (из прокамбия) и систему основных и арматурных тканей (из основной меристемы). Из прокамбия, кроме первичных проводящих тканей, образуется перицикл. По положению прокамбий и перицикл – боковые, или одну (хвощи) или несколько (покрытосеменные) так называемых инициальных клеток. Инициальные клетки меристем задерживаются на эмбриональной стадии развития в течение всей жизни растения. Именно от инициалей берут начало все латеральные (от лат. lateralis – боковой) меристемы, располагающиеся параллельно боковым поверхностям осевых органов. Вместе с основной меристемой они обусловливают первичный рост органа в толщину. У однодольных растений первичный тип роста – единственно возможный.
Вторичный рост растений, или вторичное утолщение, связано с появлением и деятельностью вторичных образовательных тканей – камбия и феллогена. Камбий может образоваться из прокамбия, перицикла или паренхимы, способной восстановить меристематическую функцию.
Как вторичная образовательная ткань, камбий дает начало вторичным проводящим тканям – вторичной ксилеме и флоэме. Феллоген, или пробковый камбий, может образоваться из клеток эпидермы или субэпидермальных клеток паренхимы первичной коры, а также из перицикла. Он дает начало перидерме – вторичной покровной ткани. Вторичные меристемы, в частности, камбий, так же как и апикальные меристемы, имеют инициали. Они несколько различаются по форме и дают начало разным элементам проводящих тканей: из паренхимотозных клеток инициалей образуются элементы паренхимы, а из прозенхимных – проводящие элементы. По положению вторичные меристемы бывают только латеральными (боковыми).
К вторичным относятся также раневые меристемы, которые образуются при повреждении тканей и органов растения. В результате паренхимные клетки вокруг раны начинают активно делиться в разных направлениях, образуется наплыв, который называется каллусом (от лат. сallus – толстая кожа, толстая мозоль). Иногда из паренхимных клеток образуется феллоген, который затягивает рану пробкой.
Цитологические признаки меристематических тканей связаны с функциями, которые они выполняют. Клетки апикальных меристем мелкие, более или менее изодиаметричные, многогранные, тесно сомкнутые, без межклетников. Они покрыты тонкой первичной оболочкой, которая содержит
Полесский государственный университет |
38 |

ОСНОВЫ БОТАНИКИ
небольшое количество целлюлозы и способна растягиваться.
Клетки меристемы характеризуются высокой метаболической активностью. В них находится густая цитоплазма с довольно крупным (относительно небольших размеров клетки) ядром. В клетках содержится много рибосом, но незначительное количество митохондрий и пластид. Эргастические вещества (вещества запаса), как правило, отсутствуют. Вакуоли мелкие, малозаметные.
Клетки меристемы имеют способность делиться в разных направлениях. Если межклеточная перегородка закладывается под прямым углом к поверхности органа, такое деление называется антиклинальным. За счет подобных делений образуются однослойные покровные ткани. Деления, при которых вновь образующиеся перегородки между клетками параллельны ближайшей поверхности органа, называются периклинальными. Они особенно характерны для латеральных меристем; за счет таких делений обычно происходит утолщение осевых органов. В том случае, если перегородка закладывается касательно окружности (тангентально), говорят о тангентальном (тангенциальном) делении. Благодаря делениям в различных направлениях верхушечная меристема стебля и корня в некоторой степени дифференцирована на слои (их можно назвать гистогенами), которые определяют функциональную дифференцировку и топографическое распределение будущих постоянных тканей.
В процессе дифференцировки клеток меристемы в постоянные ткани можно наблюдать два типа роста: симпластический и интрузивный, или интерпозиционный. При симпластическом типе рост оболочек соседних клеток происходит согласованно, их плазмодесменные связи не нарушаются. При интрузивном типе роста подобная согласованность отсутствует и одни клетки внедряются в пространство, образовавшееся между другими клетками. Он характерен для склеренхимных волокон, трахеид и т. д.
5.3 Основные (паренхимные)ткани Паренхима – это постоянная ткань, которая составляет большую часть
тела растений. Она занимает пространство между всеми другими постоянными, более специализированными тканями, ее можно обнаружить во всех органах растения. Поэтому паренхиму иногда называют выполняющей тканью. Находясь в тургесцентном состоянии, клетки паренхимы оказываются плотно упакованными, что обеспечивает дополнительную опору органам растений, особенно травянистых. При потере воды клетки спадаются и растения вянут. Паренхима – это единственная из постоянных тканей, которая может дедифференцироваться и восстанавливать меристематическую активность. Благодаря этому происходит заживление ран при повреждении растений, а также регенерация органов при вегетативном размножении. Паренхимная ткань чаще всего первична по происхождению. Она образуется из основной меристемы апексов корня и стебля. Ее клетки тонкостенные, обычно изодиаметрической, иногда вытянутой формы, стенки имеют простые
Полесский государственный университет |
39 |

ОСНОВЫ БОТАНИКИ
поры и первичное строение. Иногда клеточные стенки утолщаются и одревесневают. Паренхима хорошо развита в первичной коре стебля и корня, в сердцевине, сердцевинных лучах, мезофилле листьев, в сочных плодах.
Паренхима – единственная из постоянных тканей, которая является действительно полифункциональной. Клетки паренхимной ткани имеют различное строение и в зависимости от положения в теле растения выполняют различные функции. Независимо от положения в теле паренхима обеспечивает транспорт веществ в горизонтальном направлении по клеточным стенкам (апопласту) и цитоплазматическим путям (симпласту).
В зависимости от выполняемой функции различают ассимиляционную
(хлорофиллоносная ткань, или хлоренхима), воздухоносную (аэренхима) и запасающую паренхиму.
Ассимиляционная ткань осуществляет фотосинтез. Именно в этой ткани, несмотря на ее относительно простое строение, происходит синтез основной массы органических веществ на Земле и связывается энергия Солнца. Она состоит из тонкостенных живых клеток, содержащих хлоропласты, поэтому нередко ее называют хлоренхимой. Хлоренхима присутствует во всех зеленых частях растения. Чаще всего она залегает прямо под эпидермой. Близкое к поверхности расположение ассимиляционной ткани придает зеленую окраску органам растения. Подавляющая часть ассимиляционной ткани находится в листьях, там она составляет мезофилл (от греч. mesos – средний и phyllon – лист). Форма клеток хлоренхимы различна, соответственно им выделяют три типа хлоренхимы: столбчатую, или палисадную; губчатую, или рыхлую, и складчатую. Клетки столбчатой, или палисадной, хлоренхимы имеют таблитчатую форму, они плотно прилегают друг к другу, содержат большое число активных хлоропластов. В этой хлоренхиме интенсивно идет процесс фотосинтеза, активно осуществляется циклоз – круговое или струйчатое движение цитоплазмы. Губчатая хлоренхима состоит из более или менее округлых клеток с довольно большими межклетниками. Она расположена глубже, чем столбчатая хлоренхима, процессы фотосинтеза в ней протекают менее активно, зато хорошо развитая система межклетников обеспечивает интенсивное перемещение газов и паров воды. Дифференциацию на столбчатую и губчатую хлоренхиму можно наблюдать только у листьев, имеющих дорзовентральное строение и находящихся в условиях яркого освещения. В листьях сосны, которые обычно живут несколько лет и попадают в условия сезонных колебаний температуры и влажности, развивается складчатая хлоренхима. Она сформировалась в процессе эволюции как адаптивный признак, поскольку игольчатая форма листа резко снижает испаряющую поверхность, но должна обеспечить нормальный фотосинтез. В связи с этим в паренхиме мезофилла листьев сосны оболочки клеток врастают внутрь, образуя складки. В результате значительно увеличивается протяженность прилегающего к оболочке слоя цитоплазмы с хлоропластами и общая ассимилирующая поверхность.
Полесский государственный университет |
40 |