Рожков Ю. И. Популяции, виды, эволюция
.pdf

|
420 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
400 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
, мм |
380 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Осадки |
360 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
340 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
320 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
300 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1981 |
1982 |
1983 |
1984 |
1985 |
1986 |
1987 |
1988 |
1989 |
1990 |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
1998 |
1999 |
2000 |
2001 |
2002 |
2003 |
2004 |
2005 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Годы |
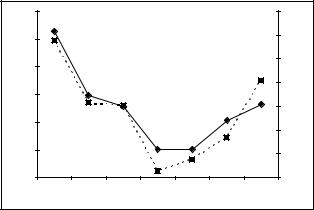
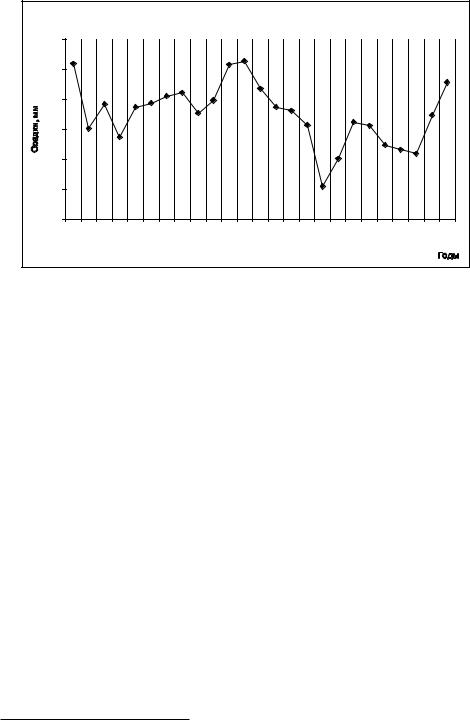
Рис. 178. Динамика среднего за 3 предыдущих года количества осадков вегетационного периода |
|||||||||||||||||||||||||
(Ломанов, 2007). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
части Азии, по-видимому, не превышала нескольких десятков тысяч человек. С такими |
|||||||||||||||||||||||||
силами наш древний предок вряд ли способен был выполнить поставленную перед |
|||||||||||||||||||||||||
ним современными исследователями задачу – истребить фауну в процессе охот. |
|||||||||||||||||||||||||
Разберем ещё одну ситуацию, которая сложилась с популяцией сайгака, обитающего в Северо-западном Прикаспии5. Обращение внимания именно на популяцию этого вида связано с рядом обстоятельств.
1.Подавляющее большинство исследователей считают, что популяция «загнана» в современную депрессию браконьерством6. А как следует из вышеизложенного, именно нерегулируемое изъятие может служить моделью охотничьей деятельности человека в плейстоцене.
2.Несмотря на относительно высокую эмбриональную плодовитость самок, по классификации В.Л. Полищука, основанной на количестве дочерей за один цикл размножения, сайгак относится к группе видов с вероятностью оказаться под угрозой вымирания 0,5 (Полищук, 2003).
3.До начала 90-х годов прошлого века пространство ареала этой популяции подвергалось негативному воздействию со стороны человека. На территории ареала обитало около 4 млн. домашних копытных, выпасавшихся на тех же пастбищах, что и сайгак. Это повлекло за собой интенсивную деградацию пастбищ. Перевыпас превратил территорию Калмыкии в антропогенную пустыню. Развиваемые пески занимали около 80% территории.
4.Впопуляциипрактическивсегдапроцветаланезаконнаядобыча.Специальнодляеё охраныещёв60-хгодахХХвекабылсозданотрядпоохране,которыйдосвоейсмертиот рук браконьеров возглавлял Улдис Кнакис. Размеры браконьерства не совсем ясны. Так, по данным Перовского (1988), доля гибели от браконьеров в общем объеме смертности
5Резкое сокращение численности населения вида отмечается и для казахстанских популяций.
6 Как принято говорить в наше время, только ленивый не писал о роли браконьерства в связи с современным состоянием популяции.
397