

Рожков Ю. И. Популяции, виды, эволюция
.pdfбразные предклеточные структуры. Они параллельно развивались в различные варианты первичных клеток, то есть с разной биохимией, генетикой, с отличающимися типами хиральности, с различными механизмами репликации, трансляции и прочего. При этом, выживали все или почти все многообразные варианты, так как в условиях низкой заселённости атмосферы жизнью конкуренция отсутствовала.
Обстоятельства изменились с падением температуры, с очищением атмосферы от «пепла» и избыточной влаги, с образованием океанов и других водоёмов. Возникли условия, в какой-то мере, подобные современным, в которых легко могли осуществляться контакты между организмами и организмов с пригодной для пищи мёртвой биологической массой, накапливавшейся в донных отложениях. Иными словами, появилась среда, способствующая эволюционной диверсификации исходных форм на ключевые звенья экосистемы. Это привело к жёсткой конкуренции. В итоге, выжили потомки только наиболее совершенной, а возможно, просто «удачливой» формы из большого числа сформировавшихся ранее в атмосфере параллельных форм. В дальнейшем эти потомки и создали всё многообразие жизни на земле.
Таким образом, падение жизни «с небес на землю», видимо, сопровождалось самой первой глобальной катастрофой – вымиранием параллельных «неудачных» форм первоклеток29.
Представленная модель предполагает первоначальную генетическую полифилетичность (сочетающуюся с исходной экологической монофилетичностью), которая после вымирания «лишних» форм заменилась генетической монофилетичностью (рис. 120.1). «Стандартная» же модель предполагает, что монофилитичность была исходным состоянием (рис. 120.2). Хотя события, развивающиеся по этой схеме, маловероятны (см. выше), в настоящее время эта модель популярна у многих исследователей,
Рис. 120. Три возможных варианта начальных этапов биологической эволюции.
1. Первоначальная генетическая полифилитичность, после вымирания «лишних» форм сменяется генетической монофилетичностью. На следующем этапе диверсификация исходной линии сочетается с отдельными актами горизонтального переноса (перемычки между ветвями).
2. Монофилитичность – исходное состояние (горизонтальный перенос символизируют на рисунке перемычки).
3. Взаимосвязанные в единую «популяцию» отдельные ветки – линии (перемычки между ветвями символизируют межлинейные «скрещивания» и горизонтальный перенос).
29 В последнее время некоторые исследователи стали «в тайне» призадумываться: «Не сохранились ли где-то на нашей планете «неудачливые» формы параллельной жизни?». Тот, кто посмелее и безответственней ответственных научных сотрудников, подхватили эти думы, призывая приступить к поиску (Дэвис, 2008).
290

что видимо является давней традицией «обязывающей» представлять эволюционный процесс как «ветвящееся дерево».
Многим, тем не менее, понятно, что объяснить зарождение жизни и начальные этапы её развития такой схемой очень сложно. Поэтому значительную популярность приобрела и другая, более усложнённая схема, кажущаяся реалистичней – эволюционный процесс исходно «стартует» как полифилетический, но отдельные стволы деревьев связаны в сеть. В результате, происходят постоянные обмены наследственной информацией между представителями отдельных линий. Обмены могут осуществляться за счёт слияний – разделений предклеток и клеток, принадлежащих разным линиям, а также за счёт передачи нуклеиновых кислот, например, в ходе процесса, напоминающего трансформацию (рис. 120.3). Весь этот «лес» эволюционирует на генетическом уровне как единое целое, что создаёт у современных исследователей иллюзию монофилетичности, которую они и изображают «стандартной» схемой (рис. 120.2).
С помощью гипотезы «связанного леса» можно попытаться объяснить становление экосистем на старте эволюции (почти в духе В.И. Вернадского) следующим образом.
Несмотря на то, что отдельные линии взаимодействуют генетически, по «пищевым» предпочтениям они вполне могут различаться друг от друга, образуя экологические цепочки. При этом, изначально плотность жизни должна быть высока, так как эволюционный процесс стартует в водоёмах. Естественно, в этой гипотезе (по сравнению с аэрозольной) появление жизни переносится на более поздний срок – на период образования водоёмов.
На первый взгляд, это выглядит совсем неплохо. Однако невозможно представить себе реальный механизм образования связей между линиями в масштабе всех водоёмов земли. А ведь только в этом единственном случае весь процесс может напоминать монофилетичный и то при условии, если обмены между линиями идут интенсивно, как бы связывая всё живое в одну сверхпопуляцию.
Сетчатая эволюция в то «первобытное» время, конечно же, не в глобальных масштабах, а в отдельных локальностях (скажем в каком-то небольшом водоёме) вполне могла происходить. Это значит, что если и не отдельные линии, то группы взаимосвязанных линий должны быть изолированы. Таким образом, мы фактически придём к чистой полифилетичности, в какой-то мере подобной изображённой на рисунке 120.1. Однако от неё надо вновь двигаться к монофилетичности, домысливая конкуренцию и вымирание всех линий (групп линий), кроме одной «избранной» (линии или группы).
Против «лесной» гипотезы свидетельствует и то, что сетчатый эволюционный процесс в действительности не стимулирует, а тормозит прогрессивную эволюцию. Чем больше взаимодействующих единиц (линий, популяций) и чем шире их ареал, тем больше торможение. Это и понятно – разнонаправленные эффекты отбора в разнородной по обширному ареалу среде взаимопогашаются при перемешивании и скрещивании особей. То же «правило гашения» действует и при случайной дифференциации (см. главы 9 и 18)30. Всё это не только легко обосновывается теоретически, но и подкрепляется палеонтологическим материалом (Буко, 1979). Нечто подобное вполне можно ожидать и для предклеток и первоклеток.
20.5. Другие взгляды на проблему происхождения жизни
Как нетрудно было заметить, все гипотетические представления о происхождении жизни имеют исключительно фантазийный характер. Несмотря на оптимизм «учё-
30 Конечно же, сетчатость и широкоареальность – не единственные, но важные причины эволюционного застоя.
291

ных», публикующихся в научно-популярной литературе, такое положение, скорее всего, будет сохраняться очень и очень долго. К сожалению, проблема происхождения жизни слишком сложна для быстрого разрешения. Однако всё же есть способ решить её «мгновенно», не «терзаясь» эволюционно-экологическими «придумками», а лишь взяв на вооружение «простые математические» рассуждения.
Попробуем ответить на вопрос: какова вероятность происхождения жизни чисто случайным образом? Время от времени такие оценки делались и давали, вроде бы, совершенно неутешительные цифры. Оценивалось появление случайным образом какого-либо функционально активного белка, присутствующего в современных организмах, и множества белков. Полученные результаты были разные, но всегда с практически нулевыми вероятностями31 от 10-130 до 10-40000. Самопроизвольное появление первоклетки в сборе,32 очевидно по ничтожности величины, не идёт ни в какое сравнение с последней из этих цифр. Обычно делается вывод, что время существования Вселенной (14 млрд. лет)33 и количество вещества, которое оно содержит, не способно обеспечить реализацию даже самой минимальной из этих вероятностей.
То есть, случайным образом ничто появиться не может.
Однако все подобные рассуждения относятся только к так называемой нашей Вселенной, которая ограничена и во времени, и в пространстве, и по заключенной в ней массе материи.
Под нашей Вселенной одни понимают потенциально видимую часть Вселенной, радиус которой измеряется в световых годах и равен её возрасту. Другие под этим термином подразумевают в несколько раз более обширную часть Вселенной. Так или иначе, космический мир, в котором мы живем, является, скорее всего, лишь частной Вселенной. Общая же Вселенная, по-видимому, безгранична и состоит из бесконечного множества частных.34 А раз это так, то нулей в цифре, стоящей в минусовой степени,
31Подробнее об этом смотрите, например, у Б.М. Медникова (2005).
32Напомним, что такая первоклетка должна содержать не только все необходимые для жизнедеятельности структуры (которые могут, в общем-то, возникнуть и закономерно за счёт самосборки
– саморганизции – см. выше), но и должна содержать некоторый минимальный для проявления жизни набор генов, оцениваемый разными авторами в 300–600 штук, и, конечно же, значительный набор белков и других соединений, необходимых для запуска биологической «машины»: механизмы репликации, транскрипции, трансляции и прочие механизмы, обеспечивающие жизнь и размножение «минимальной» клетки.
33Точнее 13,75 млрд. лет. земля же, по последним данным, появилась 4,54 млрд. лет назад (arxiv.org/abs/1001.4744). Самая последняя оценка возраста Вселенной 13,772 млрд. ± 59 млн. лет и, наконец самая, самая последняя – 13,8 млрд. ± 30 млн. лет
34Одна из наиболее популярных в настоящее время теорий, из которой следует подобное устройство Вселенной, это теория, в занимательной форме изложенная в книге Виленкина (2010) и принадлежащая группе авторов. Суть её заключается в следующем. Наша Вселенная возникла в результате инфляции (стремительного расширения пространства из микроскопической области – «трещины») сопровождаемой Большим Взрывом (результат выделения энергии при переходе физического вакуума, заполняющего «трещину», на более низкий энергетический уровень). В ходе Большого Взрыва появляется масса в виде элементарных частиц (так как энергия преобразуется в массу и обратно – E=mc2). Последующее расширение и соответственно охлаждение раскалённой Вселенной до сегодняшнего температурного состояния (подобно вырывающемуся из баллона газу, охлаждающемуся при расширении) сопровождается конденсацией вещества и образованием туманностей, звёзд, галактик и прочего. Пространство общей Вселенной постоянно «трескается» в разных местах. Одни «трещины» схлопываются, другие проходят примерно тот же путь, что и наша Вселенная или отличный от неё. Весь этот процесс идёт бесконечно во времени и пространстве, порождая бесконечное число частных (островных) Вселенных. В силу ряда физических причин, связи между частными Вселенными вряд ли возможны. В данный момент где-то «гремят» Большие Взрывы и только начинается развитие очередных островных Вселенных, а где-то частные Вселенные уже превратились в космические пустыни.
292

может быть сколько угодно. Любые, почти нулевые вероятности неизбежно будут реализованы в каких-то частных Вселенных. Конечно же, доля таких Вселенных в объёме общей Вселенной будет также практически нулевая (определяться вероятностями случайного возникновения жизни).
Во Вселенных, где произойдут подобные события, в конце концов, в ходе эволюции может появиться разумный наблюдатель. Один из этих наблюдателей, появившийся на земле во Вселенной Х (Homo sapiens), рассуждая о проблемах зарождения жизни в рамках вероятностного подхода, сделает совершенно очевидный вывод: если жизнь возникла случайно, то он (наблюдатель) во всём объёме его частной Вселенной совершенно одинок, никаких «братьев по разуму» и других «жизней» в ней не существует. В то же время в общей Вселенной и «жизней», и «братьев» бесконечное число.
Казалось бы, вот оно простое решение всей проблемы – случайное возникновение жизни и одиночество в доступной нам для наблюдения Вселенной. Однако «душа не принимает» это решение. Его так и хочется назвать «математической мистификацией». В течение нескольких столетий человечество в лице своих естествоиспытателей и философов верило и верит, что окружающий нас Космос многонаселён, а жизнь развивается закономерно на многих планетах, подобных земле. На эту тему устраиваются симпозиумы, пишутся научные книги и статьи, придумываются формулы для оценки числа обитаемых и цивилизованных планет в нашей Галактике. В зависимости от выбранных значений используемых параметров, оценки количества высокоразвитых цивилизаций варьируют в пределах от менее одной до более миллиона. живых, но нецивилизованных планет «насчитывают» значительно больше (обзор смотрите у П. Ульмшнайдера, 2009).
К сожалению, зарегистрировать сигналы искусственного происхождения, посылаемые из разных участков нашей Галактики, а тем более Вселенной, до сих пор не удалось, хотя такие попытки неоднократно проводились на протяжении многих десятков лет. На этом основании пессимисты делают следующий вывод. Либо «братьев по разуму» слишком мало, и вследствие этого, мы не можем с лёгкостью уловить идущие от них редкие сигналы, либо их много (и даже очень много), но мы им неинтересны, так как длительность их высокоцивилизованного развития десятки, сотни тысяч, миллионы и даже миллиарды лет35. Мы же развиваемся «в том же духе» несколько сотен лет, если вести отсчёт высокоцивилизованности от времени появления первых зачатков современной науки и техники, или несколько тысяч лет, если начать отсчёт от эпохи древних цивилизаций – Греции, Рима, Египта и др. Оптимисты считают, что «переразвитые» цивилизации давно находятся здесь, на земле в виде разнообразных НЛО и, следовательно, сигналить им незачем. Они полускрыто наблюдают за нами и изучают нас, не вмешиваясь в наши дела, чтобы не повредить дикарям-землянам информацией о себе и своих сверхтехнологиях, которые всё равно недоступны ни разуму, ни духовному развитию дикарей. Эта экстравагантная точка зрения имеет не только сторонников, в том числе и среди выдающихся исследователей, но и бесчисленное множество противников. Неудивительно, что среди тех, кто признаёт «тарелки» существующими, много поклонников гипотезы панспермии. Им представляется, что высокоразвитые цивилизации расселяют на них жизнь по всей Вселенной, а затем контролируют её развитие. Противники – абсолютное большинство исследователей, такие представления воспринимают как совершенно ненаучные, которые не дают ответа на главный вопрос: как первоначально появилась жизнь, если и не на земле, то на какой-то другой донорской планете, подобной зем-
35 Считается, что первые планеты земного типа появились около 10 млрд. лет назад.
293

ле? Однако умеренные поклонники гипотезы панспермии предлагают вполне научный вариант расселения жизни в пределах Солнечной системы, но не на «тарелках», а с помощью метеоритов.
Механизм этого расселения следующий. Огромный астероид диаметром в несколько километров, сталкиваясь с планетой, выбивает из неё множество осколков. Некоторые из них приобретают столь значительную скорость, что в состоянии преодолеть силу притяжения планеты. «Путешествуя» в космическом пространстве, часть осколков может достичь другой планеты и оказаться на ней в виде метеорита. Этот процесс «переброски» камней с одной планеты на другую теоретически обоснован и документально подтверждён находками на земле метеоритов марсианского происхождения (Baalke, 2005), последний из которых (а может быть и не последний) обнаружен в Марокко в 2011 году.
Остаётся вспомнить, что некоторые бактерии существуют на континентальных участках нашей планеты повсюду и даже на глубине в несколько километров, поселяясь, например, в микротрещинах. Поэтому камни – осколки – метеориты вполне могут содержать внутри себя какое-то количество транспортируемых в «законсервированном виде»36 бактерий. Каменная оболочка надёжно защищает их от жёсткого воздействия в межпланетном пространстве ультрафиолета и космических лучей. Такая защита необходима и от высоких температур при прохождении осколков через атмосферу планеты и ударе о её поверхность. Считается, что часть осколков оплавляется (за счёт трения в атмосфере и выделения тепла при ударе) лишь снаружи. Внутри же них температура может быть ниже 100 °С, что вполне достаточно для сохранения жизнеспособного состояния хотя бы некоторых бактерий, а тем более их спор37.
По мнению поклонников гипотезы «метеоритного расселения», наша земля, скорее всего, не является донорской планетой. Эту роль обычно приписывают Марсу. земля слишком массивна, и сила её гравитационного притяжения велика. Поэтому почти все выбитые из неё в ближайшее космическое пространство камни, покружив вокруг планеты, вскоре возвращаются обратно. Марс же гораздо менее массивен и, следовательно, притягивает не столь «энергично». Поэтому камни от него разлетаются по всей Солнечной системе. Кроме того считается, что 4–4,5 млрд. лет назад Марс очень напоминал землю тех же времён, но в уменьшенном масштабе. События на нём развивались ускоренными темпами и, видимо, должны были опережать аналогичные события на земле. Так почему бы не предположить, что первоначально жизнь возникла на этой планете? затем она была перенесена на землю, опередив возникновение на ней собственной жизни. Впоследствии жизнь на Марсе исчезла из-за потери почти всей атмосферы и его охлаждения38.
Эти взгляды, хотя и серьёзно обоснованы, но большинством исследователей воспринимаются как излишнее усложнение. В самом деле, зачем в общую схему происхождения жизни, которая и так проблематична, вводить ещё и дополнительные этапы
– внеземное происхождение и переселение, когда вполне можно обойтись и без них? К тому же, история науки свидетельствует: чем проще – тем лучше. То есть, сложно сконструированные гипотезы обычно отмирают, а выживают и превращаются в теории правильно объясняющие явления «простые» гипотезы. Так что лучше всего и нам
36Как известно, температуры в открытом космосе близки к абсолютному нулю.
37Бактерии выдерживают огромные, но незапредельные перегрузки, возникающие при ударе, которые ведут к разрыву клеток и тканей протист и многоклеточных. Поэтому часть из них не пострадает или серьёзно не пострадает от этого фактора.
38Но возможно оставила какие-либо следы, которые, как известно, многие годы пытаются обнаружить земляне.
294

последовать этому правилу, вернувшись к «стандартной схеме» и отбросив всевозможные современные варианты (а их много) гипотезы панспермии39.
20.6. Проблема хиральности и кода
завершая главу, очень кратко рассмотрим наиболее таинственные проблемы молекулярного устройства живого. Это, во-первых, проблема происхождения гомохиральности биомолекул – L-аминокислот в составе белков и D-сахаров40. Во-вторых, проблема генетического кода.
В течение многих десятилетий на этот счёт многократно высказывались различные, но мало что прояснявшие предположения, и только в последние годы появилась какая-то надежда на решение в будущем этих проблем. Надежда основывается на следующей оптимистической идее: «требуемая» гомохиральность появляется естественным образом по чисто физическим, а не биологическим причинам (физический фактор – гл. 15).
L- и D-изомеры только при отсутствии определённых физических воздействий выглядят совершенно равноправными. При их появлении равноправие между изомерами сразу же исчезает. «Левые» и «правые» как бы превращаются в совершенно разные по свойствам молекулы. В итоге, накапливаются в окружающей среде и вступают в реакции только «нужные» изомеры. То есть сама среда может обладать гомохирализующими свойствами, а может обладать и более «простыми» свойствами, «поддерживающими» энантиомерное равновесие. Процесс происхождения жизни был, видимо, связан именно с «гомохирализующей средой».
Как пример, приведём хорошо обоснованную гипотезу (квантово-механические расчёты) о хиральном факторе молнии (Керимбеков, 2004–2007). Принимается (вслед за В.А. Гусевым, 2001, 2002), что синтез органических соединений происходил в водных каплях первичной атмосферы под действием электромагнитного поля молнии. В качестве ключевой аминокислоты, инициирующей последующий абиогенный синтез белков, по ряду соображений, выбран L-серин. Дополнительные молекулы L- и D-серина, по-разному ориентируясь вблизи канала грозового разряда, по-разному и реагируют на его воздействие – D-изомеры разрушаются, L-изомеры накапливаются. Далее сохранившиеся и сконцентрированные в каплях L-изомеры начинают сополимеризацию, вовлекая в этот процесс L-изомеры и других аминокислот по механизму В.А. Гусева (2002). В конечном счёте, в каплях по чисто физическим причинам накапливаются L-пептиды, служащие основой для дальнейших эволюционных шагов, описанных выше.
39Иногда её ещё называют гипотезой трансспермии, имея в виду под этим «ближний» перенос жизни в «осколках» на соседние планеты. Надо отметить, что если когда-либо обнаружатся в метеоритах микроорганизмы, подобные земным, это ещё не будет доказательством возможности «переселений», так как подобные метеориты могут оказаться возвращёнными землёй собственными осколками (или метеоритами, позаимствовавшими у осколков их «микрофлору» при соударениях). Возраст подобных осколков, выбитых из нашей планеты несколько миллиардов лет назад вполне может соответствовать «стандартному» возрасту «настоящих» метеоритов (4,5 млрд. лет), так как в то время на земле, по-видимому, ещё повсеместно существовали «первозданные» породы того же возраста. Впоследствии, в ходе тектонических процессов, они исчезли. Ныне известны лишь редкие находки пород с возрастом около 3,8 млрд. лет. Все остальные породы (более 90%) значительно моложе 3 млрд. (На Луне, где отсутствует тектоника земного типа, породы сохраняют свой исходный возраст – 4,5 млрд. лет).
40Как известно, при абиогенном синтезе аминокислот их левые (L-хиральный тип) и правые (D-хиральный тип) изомеры, являющиеся как бы зеркальным отражением друг друга, встречаются с одинаковой частотой, то же справедливо и для сахаров.
295
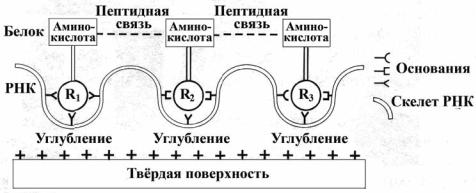
Имеются и другие гипотетические возможности, когда роль фактора, отбирающего L-изомеры аминокислот, выполняют молекулы РНК, осаждённые на поверхности твёрдого субстрата (такой субстрат в качестве ядер конденсации, как правило, включён в капли – см. выше).
Предполагается, что если цепочка РНК на субстрате подвергается электростатическому отталкиванию в периодически повторяющихся участках цепи, то это приведёт к образованию её волнообразных изгибов с углублениями, в которых могут расположиться три основания РНК. В каждое углубление помещается только одна определённая аминокислота, могущая быть (исходя из геометрических соображений) лишь L-аминокислотой. затем, между захваченными аминокислотами может произойти образование пептидной связи (рис. 121), отделение готового продукта (например, при повышении температуры) и повторение цикла (Mellersh, 1993). Эта гипотеза хороша тем, что как бы указывает на решение сразу же двух сложнейших проблем – гомохиральности и происхождения генетического кода, которые оказываются взаимосвязанными. Однако каких-то реальных подтверждений этому пока что нет.
К тому же, происходящие подобным образом события в ходе эволюции должны были каким-то образом трансформироваться в гораздо более сложные, с включением между аминокислотами и РНК посредников, напоминающих в какой-то мере ныне существующие транспортные РНК.
Так может быть не стоит «усложнять» общий ход рассуждений, а попытаться придумать гипотезу «одномоментного» с аминокислотами появления посредников? Такой путь лишь кажется упрощением, в действительности же он в ещё большей степени всё усложняет. Скорее всего, первоначально взаимодействие аминокислот и нуклеиновых кислот всё же происходило напрямую. На это «намекает» и сама структура триплетного кода.
Самые крупные аминокислоты, использующиеся в белках, не способны перекрыть более трёх нуклеотидов. (Мелкие – один, два нуклеотида).
Вырожденность кода (соответствие одной аминокислоте нескольких кодонов) как раз и коррелирует с этими цифрами (чем меньше аминокислота, тем больше вырожденность) – Керимбеков, 2004–2007. Всё это как бы указывает на то, что «предкод» в ту эпоху отнюдь не был строго триплетным – сколько аминокислота перекроет, то и было её «кодом» (для одних два нуклеотида, для других – три). В дальнейшем, с появлени-
Рис. 121. Возможный механизм сопряжённого образования триплетного генетического кода и аминокислотной L-хиральности. R1 – R3 – боковые цепи аминокислот (Mellersh, 1993).
296

ем РНК-посредников и какого-то подобия рибосом код неизбежно должен был пройти стадию унификации по максимальной своей размерности в три нуклеотида (чтобы не было потеряно аминокислотное разнообразие белков и главное – для резкого повышения определённости в их первичной структуре 41). Подобным образом возможно и развилось кодон-антикодонное взаимодействие, сохранившее память о прежнем своём предковом состоянии в явлении вырожденности.
Конечно же, всё это лишь самые общие гипотетические рассуждения. Будущее покажет, верны они или же нет.
В заключении отметим, что, несмотря на то, что в настоящее время «царствуют» L-аминокислоты, в состав пептидов клеточной стенки бактерий (пептидогликан) входят и D-аминокислоты. Известны примеры включений D-аминокислот также и в некоторые пептиды эукариот. Синтез подобных соединений никак не связан с трансляцией, а потому, видимо, и допускается использование аминокислот в D-форме. Возможно это «рудимент» той давней эпохи, когда ещё не был сделан выбор между D и L и когда совершенный аппарат транскрипции – трансляции – репликации (предполагающий гомохиральность) не существовал.
41 Полная определённость в структуре белков не достигнута и поныне. Вспомним, например, что с одного и того же гена в ходе альтернативного сплайсинга у эукариот может синтезироваться множество существенно различающихся по первичной структуре минорных белков (обычно менее 5%), по большей части совершенно бесполезных. Да и сама структура «последних» белков в разных своих частях крайне различается по «смысловой нагрузке» (одни участки крайне важны для функционирования и должны воспроизводиться точно, другие могут варьировать по аминокислотной последовательности очень широко).
297

ГЛАВА 21. СУщЕСТВУЮТ ЛИ СВЕРХПРОБЛЕМы (О ДАЛёКОМ БУДУщЕМ БИОЛОГИИ)
Не одно поколение исследователей сменится, прежде чем все малые и большие эволюционные проблемы (часть из которых рассмотрены в нашей книге) будут досконально решены. Даже глобальная и труднейшая проблема происхождения жизни, которая буквально измучила представителей всех естественных наук, в будущем, без сомнения, будет решена.
А что дальше?
Нашим потомкам-исследователям остаётся переизучение биоразнообразия (а точнее его нескончаемое переописание) и решение частных (прикладных) эколого-эволю- ционных задач?!
Эта малоинтересная перспектива (напоминающая существующее состояние географии) может длиться бесконечно.
Когда-то (на рубеже XIX – XX веков) подобные мысли посещали и физиков. Более того, в то время даже выдающиеся исследователи буквально за несколько лет до начала «Великой физической революции» (разработка теории относительности и квантовой механики), перевернувшей прежние представления об устройстве мира, заявляли, что всефизическиепроблемыразрешены(илиблизкикразрешению),ибудущимпоколениям остаётся только лишь заниматься детализацией и уточнением «преждеоткрытого»1.
Всё это происходило на фоне рассуждений философов о неисчерпаемости мира и бесконечности его познания.
Но кто слушает философов!
Один наш многоуважаемый коллега (В.С. Мирутенко) «одномоментно» объяснил нам современную трактовку этих философских представлений, изобразив на листе бумаги расширяющийся круг (в воображении конечно). Этот рисунок мы здесь и воспроизводим (рис. 122).
Всё, что внутри круга, символизирует наше «знание». Всё, что снаружи – наше «незнание», уходящее (за пределы листа бумаги) в бесконечность. Пограничная область (заштрихована) – это то, что мы и знаем, и не знаем, то есть, знаем, что это есть, но не можем объяснить. В общем, «непознанное». О существовании всего остального «запограничья» мы даже и не подозреваем. Малый круг слева – наше состояние с наукой лет 150 назад. Большой круг справа – сегодняшний день.
Очевидно, что за этот срок знаний стало много больше (круг расширился), но и количество «непознанного» столь же значительно увеличилось (пропорционально длине окружности – штриховка) и будет постоянно расти, сопровождая постоянное расширение круга. Процессу этому нет конца.
Так примерно на сегодняшний день стали рассуждать и физики, находясь, с одной стороны, под впечатлением своих достижений, с другой – сознавая свою неспособность во многих случаях что-то объяснить.
1 Удивительно, но уже в наше время (в 90-х годах прошлого века) примерно те же мысли посетили одного уважаемого всеми нобелевского лауреата, которые он и «озвучил» в одной из своих последних статей (журн. «Природа»). Буквально сразу же после этого физики заявили о новом прорыве в неведомое (о тёмной энергии и тёмной материи, в основном наполняющей Вселенную и, может быть, состоящей из частиц, подчиняющихся действию таинственных гипотетических сил).
298

Рис. 122. Объяснения в тексте.
Так где же находится биология – внутри круга и немного по его краям (а в будущем только внутри круга, как и география) или же вместе с физикой постоянно будет проникать при увеличении знаний в зону непознанного?
Попытаемся с помощью всё той же физики получить хоть какое-то подобие ответа на этот вопрос.
Теория относительности и квантовая механика, как известно, наполнены «жуткими» странностями и парадоксами. Глубинный физический смысл первой теории трудноуловим, второй же – не уловим вообще.
По общему мнению специалистов, физику квантовой механики не понимает никто, её же математика понятна всем. Поэтому частенько они же (специалисты-теоретики) высказывают мнение, что понимать физику процесса, в общем-то, и не нужно, достаточно одной математики.
Объяснение этому (уже с помощью биологов) даётся следующее. Человек с рождения (а то и до …) «изучает» окружающий мир посредством органов чувств. Мозг перерабатывает полученную информацию, обеспечивая максимальную подгонку поведения человеческого индивидуума к окружающему его миру. В итоге на подсознательном уровне формируется «жизненный опыт», на основе которого и даётся «разумное», как кажется, толкование того или иного физического явления.
Но подобная подгонка поведения у других видов, различающихся органами чувств и манерой переработки информации, вполне может привести к совсем иным представлениям об окружающем их мире. И если среди этих видов найдутся такие же великоразумные существа, как и люди, может оказаться, что их толкования физических явлений будут существенно отличаться от человеческих2.
Короче! Все эти «жизненные опыты» в значительной мере являются порождением действующего в популяциях отбора по повышению приспособленности, дивергенций, конвергенций и всего прочего, что определяет биологическую эволюцию и ей соответствует. А раз так, то «жизненные опыты» во многом видосубъективны и не могут использоваться для придания физического смысла, по крайней мере, тем явлениям, с которыми (или с подобием которых) тот или иной разумный вид (в нашем случае
– человек) никогда не сталкивался как в обыденной жизни, так и в эволюционном прошлом. То есть, то, что непосредственно не входит в сферу «жизненного опыта», не может получить представимого для человека физического объяснения. Поэтому «долой физику – ура математике».
2 Даже внутри нашего вида, по причине варьирования структур органов чувств и головного мозга, могут встречаться особи с нюансами толкования. А особи с дефектами этих структур вообще должны воспринимать физический мир как-то совсем по-особому.
299