

Рожков Ю. И. Популяции, виды, эволюция
.pdfРис. 117. Образование в атмосфере системы «капля в капле». 1 – большая капля с молекулами липидов на поверхности;
2 – испарившаяся капля, покрытая монослоем сконцентрированных на её поверхности липидов и придающих ей гидрофобные свойства; 3 – капля, обретающая вновь гидрофильные свойства после образования на её поверхности бис-
лоя липидов в ходе дальнейшего испарения; 4 – «бислойная» капля, «работая» как гидрофильное ядро конденсации, оказывается в конце концов внутри гигантской капли – водоёма;
F – показатель относительной влажности.
Прерывистым контуром выделен участок атмосферы с F < 100%.
такое ядро в итоге оказывается заключённым в другой капле, которая может превышать её по объёму в сотни, тысячи и миллионы раз. Фактически меньшая капля (ядро конденсации, гидрофильная аэрозольная «частица», липосома, везикула) оказывается как бы в водном «бассейне». Образуется система «капля в капле» (рис. 117.4).
Если мембрана не распадается, то с этого момента начинается новый период в эволюции капли – развитие системы фактически не в воздушном пространстве, а в водной среде, хотя эта среда и представляет собой водяную каплю, взвешенную в атмосфере.
Деление внутренней капли – липосомы, может происходить различными способами.
1.Мембрана разрушается («гемолиз») из-за отсутствия осмотического равновесия при чрезмерно больших размерах внешней капли. Крупная капля с объединённым содержимым разбивается на несколько мелких (механизм процесса описан выше). затем с каплями – «потомками» происходят те же события, которые изображены на рисунке 117. Из одной «родительской» капли образуются две-четыре системы «капля в капле».
2.Мембрана полностью не разрушается. Размножение внутренних капель идёт за счёт их механических разрывов от соударения облачных капель или от их раздувания в плёнку-«парашют» и в момент её разрыва (см. выше).
3.Мембрана полностью не разрушается. При попадании капель в верхние участки атмосферы с температурой значительно ниже точки замерзания воды17 образуются
верхностном слое. Начинается образование мицелл. Этот процесс описан в предыдущем разделе
– рис. 110. затем происходит объединение мицелл в ламеллы и их слияние с образующейся параллельно липидной плёнкой на поверхности.
17 Переохлаждённые водяные капли и сейчас обнаруживают на значительной высоте при температурах около -20 °С.
280
кристаллы льда, способные разорвать внутренние капли. Оттаивая в нижних частях атмосферы, «осколки» могут дать новое «потомство» внутренних капель.
4.Мембрана не разрушается. Внутренняя капля находится в осмотическом равновесии со средой. Она перемещается в атмосфере вместе с внешней каплей-бассейном, попадая в разные температурные условия. Если она окажется в сильно прогретом участке, то мембрана начнёт плавиться, её площадь увеличится при сохранении внутреннего объёма. Это приведёт к слипанию отдельных частей мембраны и делению внутренней капли на дочерние (рис. 118)18.
5.Капля, попавшая в условия пониженной влажности, начинает испаряться. С потерей воды отношение площади мембраны к объёму внутренней капли увеличивается. Это приводит практически к той же ситуации, что изображена на рисунке 118, только с учётом сокращения объёмов внешней и внутренней капель.
6.Во внутренней капле идёт активный синтез веществ, в том числе и тех, которые идут на «строительство» мембраны. Площадь мембраны пропорциональна r2, объём внутренней капли – r3 (r – радиус капли). Её объём пропорционален r2•v, где v толщина бислоя липидов. Объём внутренней капли пропорционален r2•r. Отношение двух объёмов пропорционально v/r, величине чрезвычайно малой (v << r). Учитывая, что единицей объёма капли производится некоторое количество веществ, необходимых для «наружной» и «внутренней» «застроек», то соотношение двух типов «производств» также должно быть пропорционально v/r. Иными словами, производство липидов и других соединений для мембраны может быть во много раз менее интенсивным, чем производство веществ для внутреннего пользования, так как первые используются на поверхности, а вторые во всём объёме внутренней капли. Нет особых оснований считать, что «элементы» мембраны синтезируются гораздо менее интенсивно, чем другие «строительные элементы». Отсюда вытекает вывод: рост мембраны, скорее всего, не будет согласован с ростом объёма, а будет опережать его. Вследствие этого, рост будет сопровождаться «сморщиванием» мембраны и дроблением «родительской» капли на дочернии по общей схеме, которая изображена на рисунке 118.
Испарение внешней капли может привести к образованию на поверхности мембраны слоя, напоминающего оболочку. В её состав могут входить не только вещества, «обитающие» в наружной капле, но и диффузно выделенные из внутренней капли.
Крайне интенсивное испарение может вызвать появление на поверхности протомембраны почти непроницаемого для воды кристаллического солевого слоя, который будет препятствовать дальнейшему испарению в условиях F < 100%. Если инкапсули-
Рис. 118. Механизм деления за счёт возрастания площади мембраны внутренней капли не соразмерно её внутреннему объёму (или за счёт сокращения внутреннего объёма).
18 з аметим, что действие на эукариотические клетки веществ, способствующих плавлению мембраны, приводит к отпочковыванию от поверхности множества пузырьков цитоплазмы, окружённых мембраной, и к стимуляции деления.
281
рованная внутренняя капля попадёт в условия с F > 100%, то конденсация и коагуляция облачных капель на её поверхности приведут к разбавлению и растворению солевого слоя.
Однако надо иметь в виду, что солевая капсула с жидкой каплей внутри может образоваться только при крайне малых размерах всей системы. Это обусловлено тем, что концентрации солей в облачных каплях ничтожны. Следовательно, для образования насыщенного раствора и кристаллизации требуется сверхзначительное испарение. По-видимому, более обычным должно быть полное испарение воды и из внешней, и из внутренней капель. Естественно, что при попадании такой системы-«споры» в условия с F > 100%, она будет насыщена водой, «оживёт» и начнёт вновь расти и делиться, если окажется химически и физически «препотентна».
Рассуждая на тему происхождения и эволюции предбиологических систем, различные авторы обычно делают упор на их стабильность, способность существовать, не разрушаясь, длительное время. Как правило, ничего не говорится о том, что предбиологические системы просто обязаны необыкновенно быстро разрушаться – гибнуть
– подвергаться элиминации хотя бы в силу того, что через элиминацию в значительной мере определяется действие естественного отбора. Ни для кого не секрет, что и все современные организмы «любят» быстро разрушаться. В своё время, воспользовавшись идеями Т. Мальтуса и достижениями селекционеров, Ч. Дарвин на этом свойстве построил свою знаменитую теорию «Происхождение видов путём естественного отбо-
ра» (1859).
Приведём крайне упрощённые рассуждения, иллюстрирующие это фундаментальное свойство подверженности разрушению.
Чтобы существовать неограниченно долго, любой вид должен находиться в равновесиисосредойотносительноскоростиразмноженияисмертности.Обафакторадолжны в среднем так нейтрализовать друг друга, чтобы величина популяции (опять-таки в среднем) оставалась неизменной. В качестве примера рассмотрим бактерий, которые поближе к протоклеткам, нежели другие представители биологического мира. В простейшем случае величина бактериальной популяции остаётся неизменной, если после каждого акта удвоения половина «потомства» тут же гибнет. Если учесть, что деления происходят, примерно, через каждые 20–30 минут, то количество погибших за сутки будет равно среднему числу делений и составит около 60-ти особей. Таким образом,
вусловиях постоянной быстрой гибели «потомства» (менее, чем за 20 мин.) «плата за существование» одной особи в бактериальной популяции будет равна 60-ти погибшим особям. Но эти расчёты действительны только для относительно «мягких» условий, когда наблюдается полный баланс между средой и организмом – среда понемногу, но быстро уничтожает, а организм воспроизводит, компенсируя это уничтожение. В условиях периодического дисбаланса «плата за существование» может увеличиваться до громадных размеров. Допустим, что бактерии в течение суток бесконтрольно размножаются в среде, наполненной необходимым для них ресурсом. Тогда за сутки одной особью будет произведено 260 (или иначе 1018)«потомков». Примем, что по истечении суток ресурс исчерпывается. В результате абсолютное большинство особей гибнет, а популяция возвращается к исходному состоянию, переживая «трудные времена», скажем, в виде спор. В дальнейшем, дней через 10 ресурс восстанавливается, особи оживают, начинают размножаться. цикл вновь повторяется. При таком сценарии вспышек и падений численности «плата за существование» одной особи составит 1017 особей/ сутки. Таким образом, в бактериальной популяции «плата» за жизнь каждой входящей
веё состав особи составляет от чуть меньше 102 особей/сутки до 1017 особей/сутки и более. Если рассчитать подобную «плату» для простейших, примитивных многоклеточных, позвоночных и прочих, то мы убедимся, что в эволюционном ряду при пере-
282

ходе от элементарных к более сложным организмам идёт стремительное (на порядки) падение этой величины19. Поэтому, в некотором смысле, «плату» можно использовать как показатель эволюционного прогресса организмов. Поставив в начало эволюционного ряда предбиологические системы, сразу делаем вывод, что «плата за существование» у предклеток, была никак не меньше, а скорее всего, значительно больше, чем у современных бактерий.
Только энергетическая среда может обеспечить высокую интенсивность распада протоклеток, а, следовательно, и возможность их эволюционирования. Консервирующая же среда, напротив, к подобному не способна. Поэтому все «придумки» о возникновении жизни в глубинах океана (за исключением участков, прилегающих к гидротермам), а тем более в ледяных полях или вблизи них, как иногда предполагается, можно рассматривать лишь как чистые фантазии. Для существования и эволюционного совершенствования предклеток необходим очень мощный распад предбиологических систем и столь же мощное их воспроизведение, компенсирующее распад.
Предбиологические системы возникают самопроизвольно за счёт действия физического фактора (гл. 15), который ответственен во всей неживой природе за «анатомию»
и«физиологию» объектов и различного рода структур (от вселенной до звёзд, планет, молекул и т.д.), формирующихся и эволюционирующих, согласно действию известных
инеизвестных нам физических законов.
Среди бесчисленного количества аэрозольных предбиологических систем, непрерывно создающихся и разрушающихся природой, непременно возникали и такие, у которых содержимое было более удачным для последующей эволюции. Это были системы, которые могли воспроизводить себя (по крайней мере, на уровне воспроизводства бактерий) по рассмотренной выше схеме «мира метаболизма» (рис. 114). Включившийся на этом этапе фактор отбора, конечно же, способствовал накоплению наиболее удачных вариантов, но, скорее всего, в то время он не определял ход эволюции в полной мере. Физический фактор, видимо, ещё долго был основным, лишь постепенно сдавая позиции по мере превращения неживого в живое. Он «предлагал» определенный набор возможных, с физической и химической «точек зрения», вариантов, из ограниченного числа которых отбор и «выбирал» наилучшие.20 То, как конкретно возник и формировался метаболизм, а впоследствии и системы репликации, транскрипции и трансляции, должны ответить химики. Это сверхсложная задача, видимо, будет решена лишь в отдалённом будущем.
Следует отметить, что в изложенную гипотезу происхождения жизни в атмосфере могут быть легко переделаны многие другие гипотезы, которые исходят из того, что предбиологические системы возникли и эволюционировали в других средах. В частности, мы воспользовались этим, когда иллюстрировали аэрозольную гипотезу рисунком из океанической (водоёмной) гипотезы (рис. 113). Первопричина такой «всеядности» аэрозольного сценария происхождения жизни проста. В разных слоях первобытной атмосферы фактически могли существовать любые необходимые для других сценариев условия, включая и синтез полимеров на таких вспомогательных «матрицах», как
19При таких сравнениях расчёты надо делать более точно, исходя не из количества «родителей» и «потомков», а более верным способом, исходя из их биомассы, так как размеры тех и других у высших организмов могут крайне различаться. Например, потомки у позвоночных могут существовать и гибнуть в виде клеток-зигот, эмбрионов, взрослых особей. Биомассу потомков, «использованных для платежа», следует отнормировать биомассой родителей. В результате получим показатель «платы» в эффективном числе особей в сутки (то есть выраженном в таком числе особей, когда все потомки равны родителям по массе).
20Об определяющей роли физического фактора в процессе предбиологической эволюции смотрите также у Галимова (2001).
283

кристаллы льда и минеральные частицы подходящего состава. Вспомним о насыщенности ранней атмосферы пылью, поставляющей ядра конденсации. Влажная атмосфера, пригодная для развития жизни, сформировалась заведомо раньше, чем «земная твердь» и воды на её поверхности. Если жизнь первоначально действительно появилась в атмосферном «океане», то значительно позднее она была смыта дождями на поверхность остывшей земли и в водоёмах нашла практически те же условия, что и в жидкокапельной фазе облаков. Видимо, дата обнаружения первых следов жизни на нашей планете (3,8 млрд. лет назад) отмечает минимально возможную дату «смыва». Собственно зарождение жизни в атмосфере произошло, скорее всего, около 4-х млрд. лет назад, а может быть и ранее21.
Сохранились ли в современной атмосфере реликты той эпохи? Нам кажется это маловероятным, поскольку первобытная атмосфера по всем параметрам резко отличалась от существующей. Тем не менее, и сейчас возможны «повторы» отдельных предбиологических стадий. В атмосфере (сотрудниками Американского национального управления по атмосфере и океану) были обнаружены капли органических материалов, заключённых в маслянистую оболочку, в которых на свету при наличии железа и никеля в принципе могли идти разнообразные реакции. Вполне вероятно, что эти капли возникли тем же путём, что и предбиологические. Однако произошли они явно из отходов органики биологического происхождения. Поэтому к «реликтам» они никакого отношения иметь не могут, но механизм, сформировавший их, по-видимому является «реликтовым» (рис. 117).
А что же с возможностью жизни бактерий в современной атмосфере? Неужели неизвестны те из них, которые смогли приспособиться к ныне существующей облачной среде? В своё время приводились видовые описания подобных бактерий. Но так как в применявшихся условиях культивирования, имитировавших, в какой-то мере, атмосферные условия, они делились всего лишь 1–2–3 раза, эти описания до сих пор многими микробиологами воспринимаются скептически (заварзин, 2004). Скорее всего, нет особых атмосферных бактерий. Сегодня, как и 4 млрд. лет назад, видимо процветают отдельные универсальные виды, которые могут обитать в различных, с нашей точки зрения, условиях (но, видимо, очень сходных с их «точки зрения»). Например, известно, что практически в поверхностной плёнке воды различных водоёмов обитают цианобактерии – первичные продуценты, входящие в состав нанопланктона приповерхностного водного слоя (цыбань, 1970; зайцев, 1974). Этот нанопланктон с брызгами воды постоянно вбрасывается в атмосферу и вполне может (до момента обратного попадания в водоёмы с дождями или путём оседания) продолжать там проживать и размножаться в лице отдельных своих представителей, о чём отчасти свидетельствуют и натурные наблюдения микробиологов (Кордюм, 1959;
Влодавец, 1972) (рис. 119).
То, что это, в принципе, может происходить в облачных каплях, состоящих из почти дистиллированной воды, всякий может убедиться, поставив простейший эксперимент. Для этого достаточно набрать в стеклянный сосуд воду из дистиллятора и выставить его на солнечный свет (естественно, некоторое время не закрывая пробкой). Через несколько дней или недель (в зависимости от складывающихся обстоятельств) на стенке сосуда, обращённого к свету, образуется цианобактериальная плёнка. Естественно, примерно то же вполне может иметь место и в каплях, включающих цианобактерии или, скажем, прохлорофиты. Стенки сосуда (если наличие их является необходимым условием для роста какого-то вида) вполне может заменить силикатная или какая-либо
21 Вполне возможно, что окончательное формирование первых клеток происходило так, как представляют себе авторы только что предложенной геотермальной гипотезы (Mulkidjanian et al., 2012) – в горячих лужах геотермальных полей (близь вулканов первобытной земли).
284
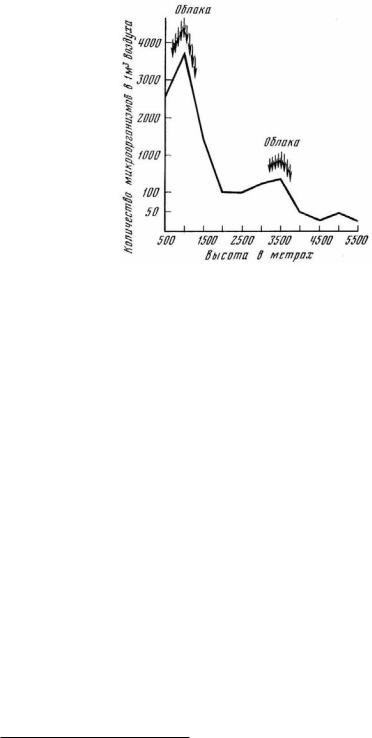
Рис. 119. Влияние облачности на содержание бактерий на различных высотах (Влодавец, 1972).
иная (подходящая для этих целей) минеральная пыль, постоянно присутствующая в современных облаках и выполняющая роль ядер конденсации – тех мельчайших частичек, на которых и формируются капли22.
20.3. Как долго продолжалась предбиологическая эволюция
Попытаемся дать ответ на следующий вопрос. В течение какого срока в ходе эволюции из предбиологической системы могла сформироваться живая клетка? Объективные суждения на этот счёт отсутствуют. Поэтому каждый пишущий или размышляющий на эту тему исследователь «придумывает» свою цифру. Диапазон этих величин составляет от тысяч до сотен миллионов лет.
На первый взгляд, кажется, что в решении этого вопроса нельзя использовать данные о темпах биологической эволюции, так как она совсем другого рода. Она не связана с созданием из элементарного предбиологического «метаболизма» базовых «устройств» (типа механизмов репликации и многого прочего), которые характеризуют любую живую клетку. Тем не менее, обратимся за подсказкой к этому привычному для нас и относительно хорошо изученному типу эволюции.
Согласно палеонтологической летописи, первые многоклеточные появились около 600 млн. лет назад. за этот срок возникло множество прогрессивных биологических «конструкций», включая человека.
Считается, что эволюция почти всегда идёт рывками: быстрые изменения за относительно небольшие промежутки времени сменяются длительным существованием в практически неизменном состоянии до следующего рывка. Рывки – это периоды нарушения равновесия со средой. В состояниях же равновесия наблюдается эволюционный застой – стазис, во время которого тоже происходит эволюция, но не в явной форме (глава 3). Предполагается, что длительность рывков отличается от застойных
22 Взвешенные в атмосфере бактерии, наряду с минеральными частицами так же играют роль ядер конденсации.
285

периодов приблизительно на два порядка. Для определённости будем считать, что они соотносятся, как 1:100 (наиболее часто принимаемое различными исследователями соотношение). Иными словами, период явных эволюционных изменений, когда появляются новые виды и новые «конструкции», в 100 раз меньше периода «благополучного» застойного существования. Отсюда можно сделать вывод, что многоклеточные активно эволюционировали на макроэволюционном уровне не 600 млн. лет, а всего 6, пребывая всё остальное время в относительном «согласии» со средой.
Далее следует вспомнить, что в основном в ходе нашей «явной» эволюции создаются только варианты одной и той же удачной «конструкции», изобретённой природой, как правило, давным-давно и переделываемой с некоторыми нюансами то в один, то в другой вид. (Принципиально не отличающиеся друг от друга и несущие, по терминологии А.Н. Северцова (1939), идиоадаптации – частные приспособления к конкретным условиям). Новые прогрессивные «конструкции», позволяющие построить совершенно иные взаимоотношения со средой, создаются крайне редко (ароморфозы А.Н. Северцова). Только они и определяют прогрессивный характер эволюции.
Для определённости опять примем, что подобные «изобретения», двигающие биологический мир к состояниям большего совершенства, происходят в 100 раз реже, чем создание различных вариантов традиционных «конструкций». Тогда, чтобы узнать, какое время затрачено на прогрессивную эволюцию, необходимо 6 млн. лет разделить на 100. Получим 60 тысяч лет. Уходя от конкретики, можно принять, что многоклеточные организмы потратили на совершенствование самих себя приблизительно 100 тысяч лет (от нескольких тысяч до нескольких сотен тысяч лет в зависимости от рассматриваемых филогенетических линий и, конечно же, от сделанных нами предположений).
Если принять, что предбиологическая эволюция на уровне предклеток началась около 4-х млрд. лет назад, а дата появления первых признаков жизни – 3,8 млрд. лет, то получим 200 млн. лет. Предположим, что и в это время ход событий в целом шёл рывками, сопровождаемыми иногда периодами активного «изобретательства». Тогда, опять же через процедуру двойного деления на 100, получим время прогрессивной эволюции, равное 20-ти тысячам лет. Естественно, эта цифра означает лишь то, что прогресс от предклеточных структур до настоящих живых клеток мог совершиться, скажем, в диапазоне от тысячи до 100 тыс. лет.
Однако, вряд ли эти прогрессивные рывки в «конструкции» были разбросаны небольшими порциями по временному отрезку в 200 млн. лет, как это имело место в эволюции многоклеточных. Скорее всего, тут просматривается лишь «одномоментное» глобальное «изобретательство», когда в самом начале процесса, вблизи точки «старта» предбиологической эволюции очень быстро по геологическим меркам произошло создание жизни.23
затем, по-видимому, многие десятки или сотни миллионов лет эта жизнь мало менялась (относительно прежнего «изобретательства»). Исходя из этого, вполне можно предположить, что искомый срок находится в тех же пределах – 1–100 тыс. лет. Естественно, подобные рассуждения крайне спекулятивны. Однако, к сожалению, это почти всё, что можно предложить на сегодняшний день. Конечно, возможно пойти и немного дальше, вспомнив об определяющей роли физического фактора. Если отбор лишь «помогал» этому фактору в выборе нужного направления развития, а само развитие, по сути, было следствием физики, которая продолжала самоорганизовывать пред-
23 Так как экологическое равновесие, стопорящее активное эволюционное развитие и обеспечивающее стазис у всех существующих в экосистемах видов, ещё полностью отсутствовало (по причине полного отсутствия настоящих экологических систем, которые должны были сформироваться лишь с появлением жизни).
286

биологические системы на более высоком уровне, то сроки могут оказаться совсем небольшими – гораздо менее 1 тыс. лет. Так или иначе, весь процесс происхождения жизни, скорее всего, протекал стремительно, без торможений на отдельных этапах.
20.4. О поли- и монофилии
Проблема происхождения и ранних этапов развития жизни содержит и ещё один важный вопрос: как структурно выглядел тогда эволюционный процесс – исходно полифилетическим или монофилетическим. Все, кто всецело находятся под влиянием экологических представлений, придерживаются первой точки зрения. Её суть предельно ясно изложена в книге Г. А. заварзина (2004). «… биосфера основывается на ряде определённых циклов, отдельные этапы которых катализируются разными функционально организмами. Поэтому В. И. Вернадский сформулировал тезис, согласно которому один вид не может существовать на земле длительно, а должно было существовать сообщество видов. Это положение исключает возможность монофилии в экосистемном контексте. Элементарная логика говорит о том, что единичный организм невозможен так же, как вечный двигатель: он не может быть одновременно и продуцентом, и деструктором. Ближе всего к такому состоянию подходят фотоавтотрофы. Днём они продуценты, а ночью деструкторы. Но для замыкания цикла им пришлось бы съесть и свои структурные компоненты и самоуничтожиться». И далее. «Возвращаясь к рассмотрению сообщества микроорганизмов, следует ещё раз напомнить, что для существования в геологическом масштабе времени это сообщество должно включать организмы, осуществляющие все трофические функции, то есть в него должны входить продуценты и деструкторы для всех компонентов продуктов и мортмассы продуцентов. В сообществе, имеющем длительное существование, должен осуществляться закон сохранения масс. Фактически, даже в реликтовых сообществах не наблюдается трофической системы монофилетического происхождения. Это заключение приходится принять как эмпирическое обобщение».
Если начальный этап эволюции рассматривать с этой точки зрения, то придётся, вслед за В.И. Вернадским, принять гипотезу вечности существования жизни в вечной Вселенной и отказаться от объяснения причин её эволюционного развития «из ничего» на отдельных планетах с благоприятными для этого условиями. Останется лишь рассуждать о распространении жизни в виде готовых бактериальных сообществ по «закоулкам» Вселенной (гипотеза панспермии Аррениуса). Такой подход автоматически освобождает и от объяснения генетической монофилии (происхождение всего живого от единого предка), так как в данном случае достаточно утверждения, что генетическое сходство всех видов – это просто состояние, данное природой. Понятно, что такие рассуждения являются «дорогой в никуда», уводящей в сторону от научной картины мира. Тем не менее, во времена В.И. Вернадского подобные умозаключения выглядели вполне «научно». Тогда, к сожалению, не было определённо известно, что не только жизнь, но даже наша Вселенная имеет свой День рождения – она появилась почти 14 миллиардов лет назад24. Поэтому, сегодня взгляды о вечности живого значимы лишь для истории науки. Однако цитаты из книги Г.А. заварзина (2004) свидетельствуют о том, что представления о первозданной экосистемности всё же не забыты. Что же касается
24 Необходимо всё же отметить, что в космологии, точнее в теоретической физике, занимающейся проблемами космологии, до сих пор появляются теории, рассматривающие нашу Вселенную как вечно существующую. Тем не менее, абсолютное большинство физиков серьёзно относятся лишь к модели, описывающей Вселенную, возникшую в результате Большого взрыва, т.е. к модели Вселенной с Днём рождения.
287

гипотезы расселения жизни по Вселенной, то в настоящее время некоторые говорят об этом, как о почти доказанном факте25. Следует всё же помнить, что экосистема – это атрибут развитой формы жизни. Тем не менее, циклика в экосистемах разного уровня, вплоть до биосферы в целом до сих пор далека от идеальной, а только выглядит таковой. Даже, казалось бы, совершенно стабильные сообщества организмов в действительности находятся в квазистабильном состоянии, постепенно меняя окружающую среду. Поэтому даже в отсутствии явных локальных или глобальных экологических катастроф, вызванных чисто физическими причинами (падение астероидов, изменение наклона оси земли и прочее), само функционирование живого на земле периодически подводит себя к скрытым «катастрофам». Это ведёт к преобразованию экосистем и, как следствие, к активации явного эволюционного процесса – завершению стазиса и началу нового рывка.
Экосистемная организация жизни должна была развиться с появлением самой жизни, но не одномоментно с ней. Обратив внимание на то, как возникает и развивается экосистема на пустом месте, можно получить подсказку, каким образом это происходило в интересующее нас время.
Первыми всегда появляются и закрепляются продуценты в виде автотрофов того или иного типа, а уже за ними поочерёдно выстраивается и вся экологическая цепочка, включая деструкторов, использующих в пищу всех её членов и замыкающих в итоге цикл. Тут действует более чем очевидное правило: сначала должна появиться пища, а затем использующий эту пищу организм.
Следуя этой простейшей логике, для запуска процесса создания первичной экосистемы было достаточно, чтобы на первобытной земле появился хотя бы один вид – автотроф. Он и мог стать основой для дальнейшей «сборки» экосистемы путём добавления других специализированных видов. Так как взять их в то время было неоткуда, то оставался единственный путь – эволюционная дифференциация исходного вида в недостающие экологические звенья.
Однако пока непонятно, что это был за автотроф. Это мог быть организм с фотолито- и/или с хемолитоавтотрофным типом энергетического обмена, то есть использующий свет и неорганику (или только неорганику в случае хемолитотрофного типа) для своей жизнедеятельности.
Вспомним, что в то время в большей части атмосферы земли царил абсолютный мрак, местами озарявшийся вспышками молний, а, следовательно, и УФ-излучения, которое вблизи канала молнии поглощалось пылью. Солнечный свет (видимый и УФ) проникал лишь в самые верхние слои атмосферы. В целом же атмосфера напоминала окрестности современных «чёрных курильщиков» на дне океана, отличаясь от них только тем, что «дым, гарь и копоть» были взвешены не в воде, а в жидкокапельной фазе. В нижних слоях атмосферы это сочеталось с высоким давлением и, соответственно, со столь же высокой температурой кипения воды.
Отсюда можно предположить, что первый организм вполне мог напоминать современных термофильных бактерий хемолитоавтотрофного типа. Но это мог быть и фототроф, использующий энергию УФ-лучей электрических разрядов и солнечного света, по гипотезе В.П. Скулачёва (1997) 26. Ещё менее возможно, что это был автотроф,
25К счастью, так говорят в основном «любители» науки, а не научные сотрудники, непосредственно занимающиеся эволюционными проблемами.
26Он предложил гипотетический механизм запасания энергии на заре становления жизни. Адениновая часть АДФ служит улавливателем квантов УФ света, за счёт чего происходит образование АТФ из АДФ и Н3PO4. В ходе дальнейшей эволюции (при уменьшении проницаемости атмосферы для УФ-излучения) произошла замена «аденинового» фотосинтеза на фотосинтез в видимом свете с использованием вначале ретиналя, а затем и хлорофилла (Скулачёв, 1997).
288

использующий энергию видимых лучей. Однако мог быть и какой-то универсальный организм, сочетающий в себе и то, и другое, и третье. Как это обычно бывает у неспециализированных форм, он всё делал не очень хорошо, а может даже и плохо. Однако в ходе последующей эволюции он «распался» на специализации и уже эти специализированные виды – потомки, «сосредоточившись» на отдельных функциях, стали выполнять их намного совершеннее.
Итак, первоначально возникшая жизнь не была экосистемной, как думают некоторые экологи, а постепенно переходила к экосистемной организации27.
Скорее всего, встречаемость в атмосфере живых клеток не была высокой, так как её самый нижний слой и поверхность земли, куда неизбежно попадали клетки, имели условия, несовместимые с жизнью. Иными словами, приповерхностный слой играл роль «элиминатора», не позволявшего жизни излишне размножаться и плотно заселять пространство. Отсюда можно заключить, что в то время жизнь вряд ли могла привести к глобальным физико-химическим изменениям на земле. Кроме того, напрашивается вывод,чтовтечениевсегопериодасуществованияжизниватмосфере,автотрофыбыли единственными её представителями, так как для образования пищевых цепей необходимы контакты живых объектов друг с другом или с мортмассой. Ни того, ни другого, скорее всего, не было. Вероятность встречи живых микронных пылинок, рассеянных в «безграничном» пространстве атмосферы, видимо, была ничтожна. Те из них, которые оседали на поверхность планеты, неизбежно погибали. В силу этих причин, экосистемы должны были начать формироваться только после охлаждения поверхности земли, когда жизнь с потоками воды переселилась из атмосферы на её поверхность.
Это переселение подсказывает решение и другой важной проблемы – факта существования генетической монофилии (единства происхождения всего живого). Кажется почти невероятным, что жизнь появилась в одном варианте с определённым типом генетического кода, определённой гомохиральностью молекул и т.д., от единственной предковой формы.
В поисках ещё одной подсказки обратимся вновь к «классическому» эволюционному процессу. Наблюдаются ли там подобные единичные появления предковых форм? Нет, всё обстоит иначе. Сразу же появляется пучок параллельных форм, эволюционирующих в одном направлении. Через какой-то период времени потомки наиболее удачливой формы начинают доминировать и вытесняют из экосистем всех или абсолютное большинство эволюционирующих параллельно с ними.
Это явление, по-видимому, характерно для большей части крупных групп животных и растений28. Вероятно, нечто подобное наблюдалось и в интересующее нас время. Многократно возникали и эволюционировали примерно в одном направлении разноо-
27 В Южной Африке на глубине около 3 км под землёй найден водоносный слой с температурой более 60 °С, населённый одним уникальным видом сульфатредуцирующих бактерий (Desulforudis audaxviator). Проживая в полном одиночестве, этот вид использует неорганические соединения, растворённые в воде, для получения энергии, фиксации углерода, азота и синтеза всех необходимых ему веществ. В данном случае перед нами наглядный пример «экосистемы в зародыше», прекрасно существующей около 20 млн. лет и фактом своего столь длительного существования, как бы документально подтверждающей возможность подобных же явлений на начальных стадиях эволюции (Chivian et al., 2008).
28 Выявлению закономерностей подобного рода, по-видимому, положил начало Л.П. Татаринов (1976, 1987). Он разработал ставшую уже классической схему «параллельной маммализации териодонтов» – независимого возникновения в шести разных группах зверозубых ящеров (териодонтов) признаков будущих млекопитающих. Лишь одна из этих групп дала начало практически всем современным млекопитающим (плацентарные и сумчатые). Четыре группы претендентов на ту же роль полностью вымерли после длительной конкуренции. Одна группа оставила только двух представителей – утконоса и ехидну.
289