

Рожков Ю. И. Популяции, виды, эволюция
.pdfмутаций хотя бы у некоторых из них достаточно велика. В результате насекомое передаст своему потомству полезный «приобретённый признак» – бактерий симбионтов с изменившимися наследственными свойствами» (Марков, 2010).
То есть, насекомые фенотипически также изменятся за счёт сохранения удачливых бактерий. Таким образом, проблему предлагается решить опять-таки вполне традиционно (но не на эукариотическом, а на прокариотическом уровне).
От себя добавим, что вполне может иметь место и ещё более простое решение (хотя столь же спекулятивное, как и предыдущее). Известно, что «заражение» симбионтами (паразитами) определённого вида может значительно изменять фенотип, а «излечение» возвращать к прежнему состоянию. Причём в качестве инфекционного агента могут выступать не только бактерии, но и вирусы (а также одноклеточные и многоклеточные эукариоты). В случае насекомых самый характерный и наиболее хорошо изученный пример – внутриклеточная симбиотическая α-протеобактерия вольбахия (Wolbachia pipientis), заражение которой приводит к кардинальным изменениям фенотипа, способа размножения и прочим эффектам (обзоры: Stouthamer et al., 1999; захаров, 1999; Горячева, 2004; Марков, захаров, 2005; 2006; Марков, 2010).
Почему бы не предположить, что нечто подобное происходит и с тлями («излечение» от каких-то бактериальных симбионтов в стрессовых ситуациях и переход за счёт этого в новую форму). Не исключено, что в некоторых случаях так всё и происходит. Однако, по нашему мнению, загадочные опыты Г.Х Шапошникова более надёжно могут быть «объяснены», исходя из ранее рассмотренных эпигенетических механизмов.
По-видимому, два близкородственных вида тлей-монофагов (D. anthrisci и D. chaerophyllina) – результат эпигенетического видообразования. Если это действительно так, то произошли они, скорее всего, от менее специализированного предка-поли- фага, который мог с равным успехом использовать оба кормовых растения (а может быть и большее их число), эпигенетически «переключаясь» на разные «образы». При последующем изменении среды (обеднение флоры) территории дифференцировались по наличию-отсутствию тех или иных кормовых растений, что и создало условия благоприятствующие появлению монофагии. Лишние «образы» в популяциях были «забыты». А дальнейшая коэволюция растения и паразитирующего на нём животного используемые «образы» – даны несколько преобразовала.
Сильнейшее стрессовое воздействие, применённое в опыте, позволило визуализировать «забытые лица», которые при сравнении их с используемыми, как и положено не вполне совпали. Визуализация происходила в течение нескольких поколений. Одни особи были успешнее в этом других.
Сильнейшая элиминация менее приспособленных (отбор) сопровождалась повышением изменчивости (за счёт появления особей с той или иной степенью выраженности забытого «образа»). Вскоре именно отбор закрепил окончательно «новый – старый» фенотип – «образ». То есть всё происходило примерно так как это изображено на рисунке 95.4–6. Однако отбор использовал не генетическую изменчивость (которая здесь отсутствовала), а эпигенетическую.
Ранее мы упустили из виду так называемое эпигенетическое наследование. Оно не связано на прямую с самим генетическим «текстом», записанным в первичной структуре молекул ДНК (то есть с последовательностью нуклеотидов) и определяемую известными (и не известными по сей день) «надгенетическими» механизмами.
Один из возможных механизмов такого рода – метилирование генов, т.е. пришивание метиловых групп (-СН3) к некоторым нуклеотидам (обычно к цитозину). Прошитые метильными группами гены теряют или резко снижают свою активность. Деметилирование, естественно, имеет обратный эффект. Подобные процессы, по-видимому, играют важную роль при индивидуальном развитии многоклеточного организма, регу-
220
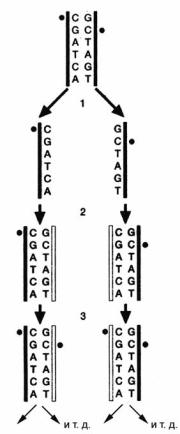
лируя (наряду с другими механизмами) активность генов в разных тканях. Важнейшую роль здесь играет метилирование (с помощью метилтрансфераз) цитозина, за которым следует гуанин (цитозин переходит в 5-метилцитозин). Такие эпигенетические модификации могут передаваться следующему поколению соматических клеток способом, изображённым на рисунке 96. То есть осуществляется фактически эпигенетическое наследование характера метилирования, а значит и характера действия (илинапротивбездействия)генов,совсемивытекающими отсюда последствиями, в линии делящихся клеток. Можно предположить, что примерно тот же механизм (или/и какой-то иной, например, связанный с модификацией гистонов и т.д.), запущенный в половых клетках (и их предшественниках), вполне может приводить к подобному же наследованию. Сейчас ряд исследователей полагает, что в некоторых редких случаях такие факты (наследование «как бы по Ламарку») уже обнаружены.
Всистеме с эпигенетическим наследованием могут происходить и своего рода «мутации» – случайные (или кажущиеся случайными) изменения в характере метилирования, происходящие на порядки величин чаще, чем обычные мутации. А также подобные же изменения под влиянием различных стимулов, что лежит в основе дифференцировки развивающегося организма, и в принципе может приводить к изменению «облика» также и отдельных особей в популяции (т.е. к различной дифференцировке особей в популяции).
Таким образом, клон вполне может подвергаться действию отбора и очень быстро эволюционировать, если он обладает описанной выше «наследственностью» и «изменчивостью».
Впоследствии, в ходе генетической ассимиляции или/и действия случайных процессов (дрейфа генов
ипрочее) в популяциях, различающихся «образами», закрепятся и настоящие генетические различия.
Вслучае с рассматривавшимися выше видами тлей «образы» скорее всего запрограммированы в ходепредыдущейэволюции,аотбор,использующий эпигенетическую изменчивость/наследственность, по-видимому, нужен лишь для того, чтобы перепрограммировать вид в целом на другой «образ», созданный в предшествующий (возможно очень отдалённый) период «классической» эволюции.
К сожалению, на данном уровне наших знаний все эти рассуждения крайне гипотетичны и даже спекулятивны, в чём можно убедиться, анализируя
Рис. 96. На этой схеме показано, каким образом может быть сохранён характер метилирования ДНК при её репликации. Метиловая группа представлена чёрным кружком. После разделения двойной спирали родительской ДНК на этапе 1 и репликации обеих цепочек ДНК на этапе 2 новые цепочки «проверяются» метилтрансферазой. Этот фермент может определить, что метиловая группа на каком-либо цитозиновом мотиве одной цепочки молекулы ДНК не соответствует только что синтезированной цепочке. Тогда метилтрансфераза присоединяет метиловую группу на новую цепочку (этап 3). Это происходит только в тех случаях, когда основания ц и Г стоят друг за другом. Благодаря этому процессу обеспечивается сохранение характера метилирования ДНК при её репликации и делении клетки (по Кэрри, 2012, с небольшими изменениями).
221

эпигенетическую литературу общего характера (Jablonka, Lamb, 1994; Эпигенетический …, 2006; Кэрн, 2012 и многие другие публикации).
19.2. Симбиогенез и происхождение эукариотической клетки9
Эукариотам в высшей степени свойственно сосуществование в рамках одного «организма» с различного рода симбионтами (паразитами) как эу-, так и прокариотной природы. Причём проживать такие эу-, прокариоты могут внутри клеток хозяина, на поверхности этих клеток и за их пределами.
Общеизвестна особая «любовь» простейших к такого рода сосуществованию с прокариотами. Некоторые из них бывают буквально нафаршированы симбиотическими бактериями (Фокин, 1993, 2011; Gőrtz, 1986)10. Но и свободноживущим бактериям, служащим пищей для простейших, свойственно вступать с ними в не менее «странные» отношения. Время от времени жертва, поглощенная протистом (рис. 97, а), может, не утилизируясь им, пережить в нём «трудное время» и даже может начать размножаться, в конечном счете, вызывая гибель хозяина (Пушкарёва, 2006).
здесь как бы реализуется своеобразная смесь двух экологических систем: хищникжертва (простейшее-бактерия) и паразит-хозяин (бактерия-простейшее)11.
От такой сдвоенной системы до чистого паразитизма один шаг. А далее невелик путь и от паразитизма до симбиоза12. Во всяком случае подобное наблюдалось в лабораторных условиях (Jeon, Jeon, 1976; Jeon, 1994, 1995 – показано, что при заражении Amoeba proteus бактериями, вызывающими массовую гибель протиста, через некоторое время между про- и эукариотом возникают симбиотические отношения).
Таким образом, внутриклеточные симбиозы не только в прошлом, но и в настоящее время возникали и возникают, по-видимому, относительно легко.
Особую роль эти эндосимбиозы сыграли на ранних этапах формирования эукариот, когда захваченные предэукариотическими клетками α-протеобактерии постепенно специализировались в митохондрии, а затем уже настоящими эукариотами захватывались фотосинтезирующие бактерии, специализировавшиеся в ходе симбиогенеза в хлоропласты (пластиды).
До сих пор некоторые исследователи не очень доверяют тому, что всё происходило именной так (через симбиоз). По крайней мере, в отношении митохондрий они пытаются придерживаться точки зрения (популярной в 70-х годах прошлого века), согласно
9Основные представления о роли симбиоза в эволюции эукариот в начале прошлого века сформулировали А.С. Фаминцын (1907) и К.С. Мережковский (1909). Самые же первые представления на этот счет появились ещё в XIX веке. Длительное время эти идеи находились в полузабытом состоянии (Хахина, 1979). В конце ХХ века они, наконец, получили общее признание
(Margulis, 1970; Маргелис, 1983; Cavalier-Smith, 1995; Тахтаджан, 1973).
10При этом бактерии могут встречаться во всех компартментах клетки (в цитоплазме, ядрах, эндоплазматическом ретикулуме и др.).
11Конечно же, для очень многих видов бактерий и простейших эти системы существуют не только в «смеси», но и в чистоте.
12Одни экологи рассматривают паразитизм, как частный случай более общего явления – симбиоза. Другие считают, что паразитизм – это самостоятельное явление. Придерживающиеся первой, по-видимому, наиболее распространённой точки зрения, иногда именуют паразитизм патогенным, антагонистическим или паразитическим симбиозом, а истинный симбиоз именуют мутуализмом (менее истинный комменсализмом, нейтрализмом и т.д.). Иногда они же говорят о симбиозе в широком смысле слова (с включением паразитизма) и в узком смысле (без включения паразитизма). Как правило, ни то, ни другое не оговаривается, а следует из контекста. Понятно, что узкий смысл первых фактически совпадает с точкой зрения вторых. Этого «совпадающего» смысла мы и придерживаемся, используя термины паразитизм и симбиоз.
222
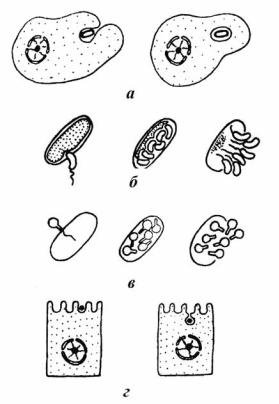
Рис. 97. Хищничество и проникновение вирусов в клетки прокариот и эукариот: а – фагоцитоз бактерии эукариотным протистом; б – нападение хищной бактерии Bdellovibrio на кишечную палочку; в – инъекция вирусной ДНК при вирусной инфекции прокариот; г – проникновение вируса в эукариотную клетку путём провокации эндоцитоза (Малахов, 2003).
которой эти органеллы развились всё же не в результате симбиогенеза, а автогенеза (то есть в результате дифференцировки в ходе эволюции отдельных компартментов предэукариотической клетки в митохондрии) – Иорданский, 2001; Чайковский, 2008. Это, однако, вопиюще противоречит генетическим данным. Кроме того, известно, что подобные симбиогенезы происходили неоднократно и независимо в разных линиях эукариот. Элементарный пример – различное происхождение пластид красных и жёлтых водорослей – рис. 98 а-г.
На поздних стадиях эволюции шло формирование и конструктивно более сложных симбиозов, в основе которых также лежит питание крупного хищника мелкой жертвой.
При большом различии в размерах первобытных (да и ныне существующих) протистов в качестве жертв использовались (и используются) не только бактерии, но и мелкие формы эукариот. Если последние уже сформировали симбиотические отношения с бактериальными клетками, а первые ещё нет, то такие мелкие эукариоты-жертвы могут быть «пощажены» хищником, и в ходе дальнейшего симбиогенеза вполне может сформироваться облигатная (постоянно существующая) симбиотическая «матрёшка»: бывшая бактерия – бывший мелкий эукариот – крупный эукариот. Подобное устройство имеют криптофитовые (Cryptophyta) и некоторые другие водоросли (Малахов, 2003, 2004) – рис. 98 д-и. Общеизвестны и факультативные «матрёшки» (результат
223
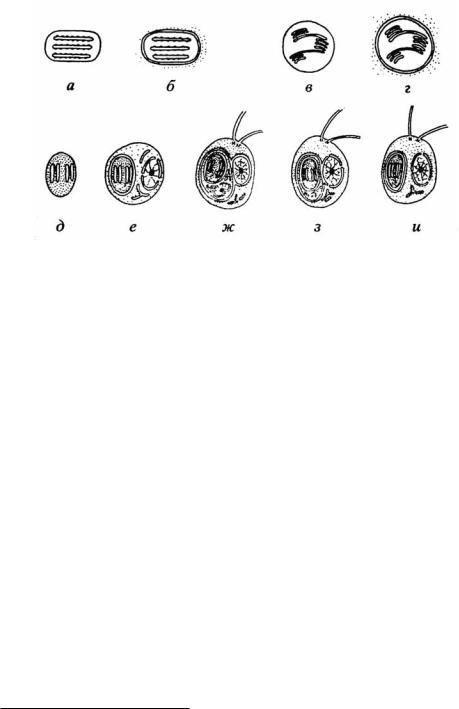
Рис. 98. Пути происхождения пластид у эукариот: а, б – происхождение пластид красных водорослей; а – свободноживущая цианобактерия с хлорофиллом а и фикобиллином; б – пластида красных водорослей, в, г – происхождение пластид зелёных водорослей и высших растений; в
– свободноживущий прокариот с хлорофиллом а и b, г – пластида зелёных водорослей,
д-ж – происхождение пластид криптомонад, хризоманад и динофлагеллят; д – гипотетический свободноживущий прокариот с хлорофиллами а и с,е – эукариот, имеющий прокариотного симбионта с хлорофиллами а и с, ж – криптомонада, имеющая эукариотного симбионта с рудиментарным ядром – нуклеоморфом, з – редукция нуклеоморфа у хризомонад, и – редукция самой наружной мембраны в пластидах динофлагеллят. Точки вокруг пластид на фрагментах б и г символизируют внутриклеточную среду (Малахов, 2003).
ещё невполне завершённого, как у криптофитовых, симбиогенеза). Например, парамеции Paramecium bursaria с включёнными в них зелёными водорослями (хлорелла). Те и другие способны существовать и по отдельности (но менее успешно, чем в «содружестве»)13.
Одни и те же или относительно близкие виды бактерий и, к примеру, упоминавшиеся выше зелёные водоросли могут быть симбионтами самых различных групп одно- и многоклеточных эукариотов. Нет никаких оснований предполагать, что в далёком прошлом ситуация чем-то существенно отличалась от нынешней. В том числе и в те времена, когда из эндосимбионтов формировались органеллы. Так что, скорее всего, события, приведшие к образованию органелл бактериальной природы, в начальный период симбиогенеза должны были происходить так, как это изображено на рисунке 99.2, а вовсе не так, как показано на рисунке 99.1, и как это обычно принимается. То есть в разных филогенетических линиях эукариот органеллы могут в общем-то принадлежать к разным линиям прокариот. Однако так как участвовавшие в этих событиях прокариотные и эукариотные виды входили, видимо, в состав филогенетически родственных групп, никакими генетическими методами (позволяющими дифференцировать путём построения дендрограмм лишь достаточно удалённые друг от друга группировки – см. гл. 3) невозможно отличить один тип событий от другого14.
13з аметим, что в прошлом бывали случаи, когда симбионт-содержащую форму какого-либо вида и по сути ту же форму, но без симбионтов описывали как два разных вида.
14Отметим, что исходную симбиотическую полифилию предполагал ещё в начале ХХ века один из создателей концепции симбиогенеза (и самого этого термина) К.С. Мережковский.
224
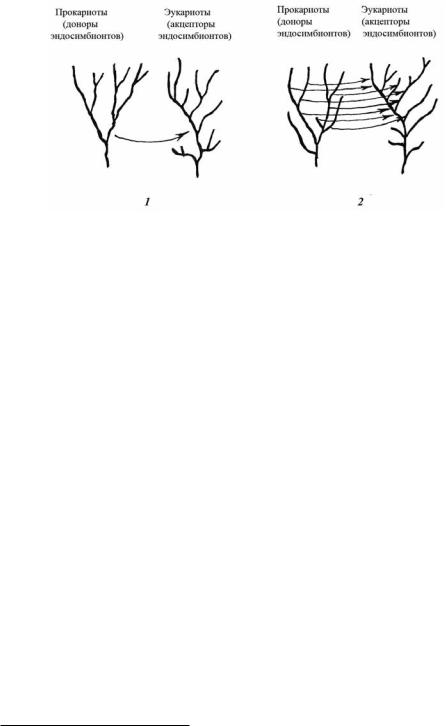
Рис. 99. Гипотетические филогенетические деревья для монофилетического (1) и полифилетического (2) симбиогенезов. Стрелки, отражающие передачу симбионтов от про- к эукариотам, и деревья выполнены в произвольной форме.
Только пластидами и митохондриями симбиогенез, приведший к созданию этих органелл, видимо не ограничивается.
Как мы только что отмечали, в бактериальном происхождении пластид не сомневается практически никто. Происхождение митохондрий тем же путём вызывает сомнение у очень незначительного числа исследователей, а вот в отношении того, что жгутики и реснички (ундулиподии) эукариот – это «потомки» спирохетоподобных бактерий, сомнения возникают до сих пор у многих.
Тем не менее, известны симбиотические системы, как бы иллюстрирующие те древнейшие события, которые возможно привели к «приручению» предками эукариот подвижных бактерий, напоминающих ныне существующих спирохет. Это жгутиконосец
Myxotricha paradoxa, обитающий в кишечнике термита Mastotermes darwinensis.
Поверхность его покрыта несколькими сотнями тысяч спирохет-экзосимбионтов, которые, осуществляя согласованные движения на манер настоящих ресничек, перемещают хозяина в пространстве кишечника.
Этот случай Лин Маргелис использовала для иллюстрации своей гипотезы симбиотического происхождения ундулиподий (Маргелис, 1983)15.
По современным представлениям, молекулярная структура ресничек/жгутиков не соответствует таковой спирохет. Так что бактериальные предки первых (если они были), скорее всего, неродственны предкам вторых. Однако это вовсе не означает, что в перспективе не будут обнаружены бактерии-«ундулиподии». Вспомним, что длительное время свободноживущие бактерии с хлорофиллами «a» и «b» также не были известны. Поэтому пластиды зелёных водорослей и высших растений производили от цианобактерий, содержащих хлорофилл «a». Специфические для них красные и синие пигменты – фикобилипротеины, впоследствии (на стадии симбиоза), как предполагалось, могли быть утрачены, а хлорофилл «b» мог быть приобретён в ходе дальнейшей эволюции. Не исключено, что так всё и происходило. Однако в 70–80-х годах прошлого века была наконец обнаружена группа фотосинтезирующих бактерий – прохлорофитов, содержащих хлорофиллы «a», «b». Сейчас считается, что вероятнее всего далёкие
15 Справедливости ради надо отметить, что ещё Б.Н. Козо-Полянский (1924) предполагал, что центриоли являются по всей видимости одним из дериватов жгутиков (точнее их базальных телец – кинетосом) и имеют бактериальную природу.
225

предки именно этих бактерий могли являться и предками хлоропластов высших растений16.
Итак, с учётом того, что отдалённый родственник первобытной симбиотической бактерии-«ундулиподии» когда-нибудь найдётся (как в случае с прохлорофитами), мы присоединяемся к тем, кто верит в симбиотическое происхождение жгутиков. К тому же внутри базального тельца жгутика (кинетосомы) находится крохотная кольцевая молекула ДНК, содержащая всего несколько генов (видимо участвующих в образовании кинетосомы). Эту молекулу вполне можно рассматривать, как реликт бактериальной хромосомы.
Общая тенденция, характерная для всех симбионтов – это упрощение организации и сокращение числа генов. Обычные симбиотические бактерии содержат их раза в два меньше, чем свободноживущие виды (3000–4000 генов с колебаниями в ту и другую сторону), а вот митохондрии (в зависимости от вида, в котором они «проживают») имеют их обычно лишь до нескольких десятков (в среднем 40–50 генов). Хлоропласты зелёных водорослей и высших растений (сформировавшиеся значительно позднее митохондрий) – где-то около 120 генов.
Гены симбионтов либо теряются за ненадобностью при специализации, либо «перекочёвывают» в геном хозяина и оттуда «руководят» (или не «руководят», находясь в инактивированном состоянии), либо используются хозяйские гены (как находящиеся в его геноме, так и, в свою очередь, «перекочевавшие» в геном органеллы)17.
Вектор «перекочёвки» всегда сохраняет главным образом одно направление – от симбионта к хозяину (за исключением одного генетического случая, когда симбионтом является ядро, см. ниже).
В общем, вырисовывается следующая тенденция: чем древнее связь между симбионтом и хозяином, тем меньшее число генов (и своих, и чужих) несёт симбионт. (Число генов убывает в ряду: свободноживущие бактерии, эндосимбионты, хлоропласты, митохондрии, ундулиподии).
Обусловлено это, по-видимому, одним – специализацией, позволяющей эффективней расходовать энергетические ресурсы.
Напрашивается аналогия с человеческим обществом, в котором различные производства разделены функционально и территориально. Такая структура наиболее экономически (= энергетически) выгодна. Легко себе представить, что было бы, если вдруг на каждом предприятии в одночасье решили и сталь варить, и пироги печь, и электронику собирать. Экономика, скорее всего, рухнула бы. Допустимо было нечто подобное (и то отчасти) лишь в исторически первобытную эпоху, когда по сути дела настоящего производства ещё и не было. Так и в клетке, каждый симбионт, «стремившийся стать» органеллой, должен был совершенствовать преимущественно одно какое-то «производство», наиболее важное для выполнения конкретной симбиотической функции, а от остальных (которые эффективней осуществит хозяин или другой симбионт) избавляться.
Генам же лучше пребывать в ядре – «производственной» структуре, специализирующейся на хранении и переработке информации.
Повышение уровня специализации происходило в основном за счёт отбора (конечно же, нельзя забывать и фактор выбора «профессии», см. гл. 12), действовавшего в популяциях суперорганизмов (симбионты – клетка-хозяин). Наиболее успешными (приспособленными) оказывались (как и в человеческом обществе) специализированные
16Некоторые исследователи в этом сомневаются.
17Ожидается, что минимальное число белков, которое должна содержать митохондрия, превышает шесть сотен. Из них почти все (за исключением десятка, двух или нескольких десятков) кодируются ядерными генами (Gabaldon et al., 2004).
226
по «производствам» комплексы, способные произвести больше себе подобных за ту же энергетическую цену.
Итак, симбионты, следуя «правилу специализации»,должныпостепеннопередаватьсвои генетические функции ядру клетки. Имеется ли предел этому процессу? Никаких теоретических обоснований для такого предела нет. В общем-то, вполне можно себе представить, как некий симбионт, превратившись в органеллу с ограниченным числом генов, в конце концов окажется и органеллой без генов. Последние нужные гены просто «переберутся» в ядро. При этом «тело» органеллы останется на своём месте за пределами ядра (выполнять предназначенную ему функцию), а его «душа» (информация о «теле» в виде генов) «переселится» туда, где её пребывание уместней (в ядро). Происходили ли подобные события полного «переселения» в истории эукариот и их симбионтов
– неизвестно. Но почему-то хочется верить, что происходили. Только вот выявить их мы вряд ли когда-нибудь сможем, так как полное «переселение души» органеллы формально неотличимо от результатов межвидовых горизонтальных переносов генов (см. гл. 10).
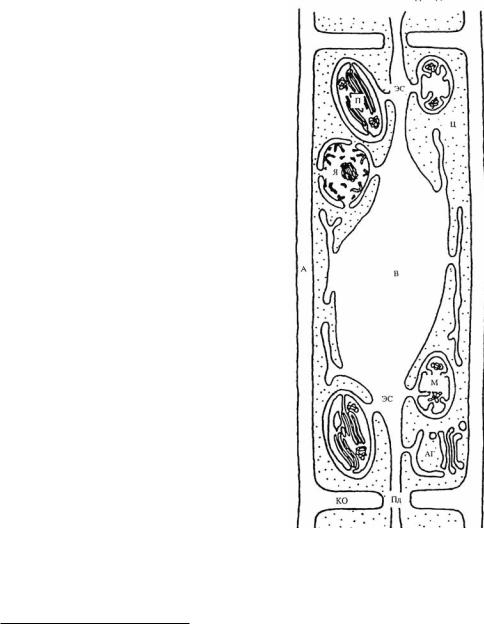
Обратите внимание на рисунок 100, изображающий растительную клетку в интерпретации Ю.В. Гамалеи (1997)18. На этом рисунке ядро как бы «уравнено в правах» с митохондриями и пластидами и выглядит вполне как органелла, окружённая двойной мембраной. Видимо созерцание подобных изображений уже после того, как эндосимбиотическая теория (в результате исследований Л. Маргелис) стала в научных кругах сверхпопулярной, некоторых привело к мысли, что
иядро появилось в результате симбиогенеза. Сразу отметим, что поклонников этой идеи
очень мало. Пока делаются только попытки
Рис. 100. Модель растительной клетки по Ю.В. Гамалею (1997): ЭС – эндоплазматическая сеть, П – пластида, ц – цитоплазма, А – апопласт, В – центральная вакуоль, М – митохондрия, Я – ядро, АГ
– аппарат Гольджи.
18 Мы приводим этот рисунок потому, что он чрезвычайно прост в отличие от многих других и главное отражает симпатичные нам идеи автора. Ю.В. Гамалей (и его последователи) считают, что наружная мембрана пластид и митохондрий (по крайней мере, у растений) вовсе не вакуолярная, а эндоплазматическая. То есть многие митохондрии и хлоропласты «проживают» не в цитоплазме (где и положено быть вакуолям), а в эндоплазматическом ретикулуме (сети), перемещаясь по клетке внутри его каналов и сообщаясь через плазмодесмы с другими клетками. Как в настоящее время выясняется, реальная картина немного посложнее (Великанов и др., 2012). Однако в нашем случае эти детали не играют существенной роли. Кстати, широкие межклеточные каналы, способные видимо пропускать крупные «предметы» (вирусы, органеллы и возможно бактерии), как недавно выяснилось, иногда свойственны и животным клеткам. Наряду с широкими имеются и мельчайшие каналы, пропускающие только молекулы.
227

формулирования гипотез19. Стандартный же подход возникновения ядра (который признают почти за истину многие) обходится вовсе без каких-либо симбиозов (рис. 101).
И всё же симбиогенного происхождения ядра никак нельзя исключать. Вспомним, что когда-то и общепризнанная сегодня эндосимбиотическая теория происхождения хлоропластов практически не имела сторонников. Как знать, может быть эта история вновь повторится. Поэтому рассмотрим очень кратко простейший сценарий эндосимбиотического появления ядерной структуры у эукариот.
Ничтонемешаетнампредположить(соглядкойнаустройствосовременногомикромира), что 1,5–2, а возможно и много более млрд. лет назад (Тимофеев, 1982; Brocks et al., 1999; Розанов, 2003) существовали достаточно крупные прокариоты20, способные вести хищнический образ жизни на манер многих современных протистов, то есть питающихся своими более мелкими «собратьями».
Такие хищники должны были иметь множество кольцевых хромосом (а не одну единственную) просто потому, чтобы обеспечить возросший объём клетки белками, необходимыми для строительства «тела» и его функционирования. «Производствен-
Рис. 101. Один из вероятных путей происхождения ядра: а – гипотетический прокариот с акти- ново-миозиновой системой, б 1 – полиплоидный прокариот, в – эукариот с ядерной оболочкой, но прокариотной организацией наследственного материала (мезокариотное ядро). Обозначения: 1 – кольцевая молекула ДНК, 2 – актиновые филаменты, 3 – фагоцитируемые пищевые частицы, 4 – глубокие инвагинации цитоплазматической мембраны, формирующие ядерную оболочку
(Малахов, 2003).
19Одна из последних предложена Г.Г.Гончиковым (2010). Основывается на отдалённом сходстве проспоры (несозревшая спора внутри клетки – спорангия) некоторых спорообразующих бактерий с ядерной клеткой. Предполагается, что посредством фагоцитозоподобного механизма обособления проспоры, лишённый клеточной стенки спорангий (L-форма) древней клостридии (имеющей и поныне эндоскелетные микротрубчатые придатки) поглотил клетку эуриархеи. В результате образовалась клеточная система «клетка в клетке». Спорангий представлял внешнюю клетку, эуриархея – внутреннюю (ядро). Микротрубчатые придатки впоследствии превратились в митотический аппарат. Другая интересная гипотеза – одновременное происхождение ядра и митохондрий через синтрофизм (объединение бактерий различных видов в колонию для питания общим для всех субстратом). центральная архебактериальная клетка колонии в ходе совершенствования сообщества постепенно приобрела функцию ядра, а переферийные клетки – функции митохондрий (Sclosse, 2003). Небольшой обзор других представлений можно найти у Г.Г.Гончикова.
20Типичные размеры современных бактерий 0,5–3–10 мкм. Встречаются также «гиганты» (до 50,
100и более 200 мкм – удлинённые и нитчатые формы) и «карлики» – 0,1–0,5 мкм. Типичные размеры современных одноклеточных эукариот 10–100 мкм. Виды «гиганты» – 1–3 мм; «карлики» – 2–10 мкм.
228

ных мощностей» одной хромосомы и ныне едва хватает лишь на то, чтобы содержать объём «стандартной» бактериальной клетки (например, такой, как Escherichia coli)21. Процесс размножения такого прокариотного хищника, скорее всего, должен был происходить, примерно так, как это имеет место быть у крупных форм микоплазм22 – фрагментация их тел и появление в итоге мелких «потомков», которые должны ещё дорасти до размеров своих «родителей» (рис. 102.1, 2).
Так как «потомки» на какое-то время перешли в размерный класс жертвы, они могут быть поглощены другим хищником, подобным их «родителю» – рис. 102.3. Этот хищник может даже принадлежать к одному виду с «родителем», а может быть и совсем другим видом. Допустим, что поглощенному «потомку» посчастливилось – он избежал утилизации, прижился и набрал необходимую для размножения массу за счёт внешней клетки23. В этом случае размеры подобного составленного из двух клеток существа увеличатся двукратно (рис. 102.4), что означает появление суперхищника – «монстра», способного использовать для питания много более широкий спектр жертв (и/ или частиц органики) и даже возможно питаться не особо крупными «родителями». При наличии соответствующей экологической ниши такое существо могло процветать и совершенствоваться в ходе эволюции24.
Процесс его размножения, затрагивающий внешнюю и внутреннюю клетку, поначалу, конечно же, должен был быть далёким от совершенства. Могли появляться и «монстры» с разным набором «ядер», и даже клетки, лишённые «ядер» (т.е. прежняя форма хищника) – рис. 102.5. Можно предположить, что в то далёкое время один из хищников, обозначенный на рисунке 102 под цифрой 3, уже нёс на себе симбиотических спирохетоподоных бактерий. (Очевидно, что активные и быстрые перемещения в пространстве важны для существ, ведущих такой образ жизни). Если симбиоз зашёл достаточно далеко, то внутри хозяйской клетки вполне могли находиться и
21Как известно, инфузории, для которых эта проблема стоит особенно остро (так как именно среди них встречаются наиболее крупные формы), разрешают её следующим образом. Содержат репродуктивное ядро (микронуклеус), которое большую часть жизненного цикла неактивно (используется в основном в половом процессе), и вегетативное (макронуклеус), функционирующее в клетке «на полную мощность». Все хромосомы в макронуклеусе по сравнению с маленьким ядром умножены в 100–500 и более раз (в зависимости от вида и объёма обслуживаемой биомассы).
22Паразитические и свободноживущие примитивные формы бактерий представляют собой, по-видимому, сборную группу. Имеют по сравнению с продвинутыми бактериями пониженное количество генов в кольцевой хромосоме. Лишены клеточной стенки и по этим причинам очень напоминают гипотетических древних прокариот. В этой связи наиболее интересна свободноживущая бактерия Thermoplasma acidophilum, морфологически и цитологически (отсутствие
клеточной стенки и прочее) сходная с микоплазмами (ранее таковой многими и считалась). Облигатный термофил (оптимум роста + 59 °С при кислотности среды рН 1–2). Выделена в 1970 г. из отвалов каменного угля, подверженного саморазогреванию. Данные по филогенетическому анализу с использованием 16 S pPНК свидетельствуют, что это вероятнее всего примитивная архебактерия. Геном её значительно меньше, чем у «типичной» бактерии (в три раза меньше, чем у «стандартной» бактерии E. coli). Все эти характеристики (архебактериальность, а значит «родственность» эукариотам; небольшой геном; отсутствие клеточной стенки) позволяют считать, что её далёкие предки могли быть одновременно и предками современных эукариот (во всяком случае, так считает ряд современных исследователей).
23Подобное «поведение» свойственно современным мелким формам микоплазм, многие из которых – это внутриклеточные паразиты, комменсалы и т.д. (т.е. эндосимбионты в широком смысле).
24В наше время свободноживущие «монстры» с «ядром»-бактерией вряд ли где-то существуют. Однако среди паразитов нечто подобное всё же обнаружено. Известна бактерия, обитающая в мучнистом червеце, в которой в свою очередь обитает другая бактерия (Von Dohlen et al., 2001). На этой основе была даже предложена симбиотическая модель ядра (Martin, Russel, 2003).
229