

Рожков Ю. И. Популяции, виды, эволюция
.pdfКроме рассмотренного выше дрейфа, оперирующего особями (зиготами, гаметами), по-видимому, может осуществляться и дрейф, затрагивающий целые группировки. В главе 3 мы уже встречались с одним из его вариантов – «филогенетическим» дрейфом, возможно действующим на макроэволюционном уровне и оперирующим видами
(Stanley, 1979).
В заключении кратко остановимся на взаимодействии дрейфа генов и случайного процесса несколько иного рода, идущего на молекулярном уровне. Это взаимодействие относится к событиям происходящим, например, внутри мультигенных семейств
–тандемно повторяющихся тесно сцепленных в хромосоме генов, родственных по структуре и функции7.
Одни мультигенные семейства включают всего несколько или один-два десятка генов, другие – многие десятки, сотни и тысячи.
Несмотря на подобные численные, а также функциональные различия между семействами, всех их объединяет одно – гены, входящие в мультигенное семейство, эволюционируют согласованно, оказываясь более сходными друг с другом, чем если бы они эволюционировали независимо. Как только это выяснилось, тут же был найден и механизм, объясняющий подобное явление – гомологичный неравный кроссинговер в сочетании со «случаем» (Smith, 1974; Black, Gibson, 1974; Ohta, 1979).
На рисунке 48 он проиллюстрирован на элементарном примере внутрихромосомного (т.е. происходящего между сестринскими хроматидами) неравного кроссинговера8, который, повторяясь из поколения в поколение, может, в конце концов, привести к «захвату» одним из вариантов гена всего семейства.
Так как такие «захваты» в «родословных» разных хромосом «по воле случая» могут осуществлять разные гены, то параллельно идёт наработка множества вариантов такого семейства в популяции. Но популяции конечны, и в них, в свою очередь, идёт дрейф генов (в данном случае в форме дрейфа вариантов мультигенного семейства). А дрейф, как известно, вполне может привести к фиксации одного из вариантов. Этот механизм, конечно же, предполагает и действие «следящего» за всем процессом отбора, отметающего параллельно образующиеся в ходе неравного кроссинговера и явно «патологические» варианты семейства (со слишком большим или слишком малым числом генов
–на рисунке 48 не показаны, но читатель без труда может их домыслить).
Кроме того, тот же отбор стремится сохранить и индивидуальность каких-то генов в семействе (обычно малые семейства), когда это необходимо для осуществления функций. В результате уровень сходства между генами семейства будет определяться совокупным действием всех факторов, участвующих в данном процессе.
Общеизвестно, что геномы различных видов насыщены тандемными повторами различной длины, не несущими в отличие от мультигенных семейств явной функциональной нагрузки, и следовательно не особенно «опекаемых» отбором (или «неопекаемых» вообще?!). здесь-то и проявляет себя в полной мере рассмотренный механизм, и сходство может переходить в идентичность всех входящих в блок повторов.
В случае многих малых мультигенных семейств, когда каждый из генов и его доза значимы, неравный кроссинговер, скорее всего, вряд ли играет какую-либо заметную роль (или не играет никакую).9 здесь те же функции гомогенизации последовательностей (также осуществляемой при поддержке дрейфа и контроле отбора) берёт на себя
7 В мультигенные семейства могут, конечно же, входить функционально неактивные гены (псевдогены).
8Результат ошибочного (со сдвигом) спаривания почти идентичных генов.
9 Кроме тех редчайших случаев, когда в ходе эволюции появляется необходимость увеличить или уменьшить число копий.
110
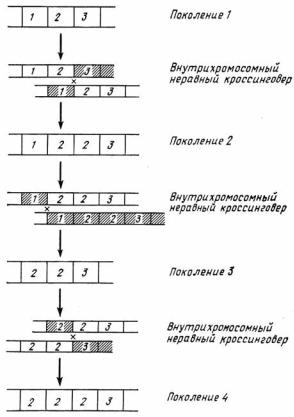
Рис. 48. Диаграмма, иллюстрирующая процесс согласованной эволюции, происходящей с помощью неравного кроссинговера, который приводит к случайной фиксации гена 2 в соответствующем участке хромосомы (Кимура, 1985).
такой «маловредоносный» способ генетической рекомбинации, как генная конверсия, которая не приводит к увеличению или уменьшению числа генов в мультигенном семействе. Но об этом мы поговорим в главе 11. А в главе 19.4 вновь вернёмся к самым общим проблемам дрейфа, затронутым в начале этой главы. Рассмотрим «дрейф по пространству», занимаемый популяционными группировками, а также разберём особенности прохождения популяциями «бутылочного горлышка».
111
ГЛАВА 7. ПОСЛЕДОВАТЕЛьНОСТь МУТАцИй И ПОПУТНый ТРАНСПОРТ
В этой небольшой главе мы очень кратко рассмотрим два эволюционных фактора, которым обычно не уделяют внимания, а часто и не упоминают, считая, что их влияние на эволюционный процесс второстепенно, а то и третьестепенно. Тем не менее, каково бы ни было их действительное влияние (малое или значительное), очевидно, что учитывать необходимо все факторы независимо от того, определяют ли они ход эволюции или всего лишь подправляют его.
Оба фактора объединяет то, что они относятся к категории случайных процессов, запускаемых в действие отбором.
7.1. Последовательность мутаций
Допустим, по какому-то количественному признаку идёт отбор. В популяции последовательно появляются мутации, которые подхватываются этим отбором. Допустим далее, что первичная популяция в силу каких-то обстоятельств распалась на две дочерние изолированные популяции. Будут ли они в таком случае дифференцироваться друг от друга? Фенотипически нет, так как направление и характер действия отбора остались прежними, и, следовательно, фенотипические изменения, по крайней мере первое время, будут происходить параллельно. Генетически, однако, популяции начнут дифференцироваться сразу же после их разделения.
Действительно, близкий фенотипический эффект может быть получен множеством различных способов на молекулярном уровне. Например, активность фермента, контролирующего какую-либо реакцию (а за ней и морфологический признак), может быть изменена сходным образом путём самых различных замещений аминокислотных остатков в его первичной структуре. Если опуститься до гена, контролирующего структуру фермента, то, учитывая вырожденность кода, число замен оснований, дающих один и тот же фенотипический эффект, должно быть ещё больше. Таким образом, существует множество самых различных мутаций генов с идентичным фенотипическим проявлением. Вероятность того, что в обеих популяциях произойдёт одна и та же мутация (из огромного набора возможных), ничтожна. Отсюда мы можем принять, что в разных популяциях под действием отбора будут фиксироваться и разные мутации (но, естественно, со сходным фенотипическим проявлением). Так как последовательности возникающих и закрепляющихся в популяциях мутаций различны, то чем дольше будет действовать отбор, тем дальше будут удаляться друг от друга популяции на уровне генотипов, сохраняя между тем определённое сходство фенотипов. По-видимому, этот эффект впервые описал Э. Майр (см., например, Майр, 1974).
Этот процесс был изучен с применением имитационных компьютерных моделей (Mani, Clarke, 1990). С помощью генератора случайных чисел осуществлялся последовательный выбор благоприятных мутаций генов, контролирующих условные количественные признаки, из определённого набора мутантных аллелей. Поскольку при этом в разных популяциях «возникали» разные мутации, популяции дивергировали, даже если их начальные состояния и давления отбора были одинаковы. Выяснилось, что в случае больших величин популяций и/или сильного давления отбора последовательность мутаций оказывается гораздо более эффективно дифференцирующей популяции силой, чем генетический дрейф, также представляющий собой случайный процесс, но действующий в прямо противоположных условиях – при малых размерах популяций и слабом давлении отбора (или его отсутствии – гл. 6).
112

7.2. Попутный транспорт
Нейтральный ген может быть подвержен действию косвенного отбора за счет сцепления с «адаптивным» геном, по которому и идет отбор. Этот попутный транспорт (Maynard Smith, Haigh, 1974) относительно недолговечен – от нескольких поколений до десятков поколений (в зависимости от близости сцепленных локусов). Рано или поздно связи между генами в процессе кроссинговера изменятся так, что нейтральный аллель, частота которого ранее росла, в конце концов окажется «заурядным», а выделится другой нейтральный аллель из того же локуса, и точно так же попутным транспортом начнет наращивать частоту на фоне падения частоты альтернативного аллеля, и т.д.
Так как осуществление кроссинговера по длине хромосомы случайно1, и, кроме того, различные аллели нейтрального гена случайным образом удачно «цепляют» время от времени удаленные от них на различные расстояния «адаптивные» гены, то динамика нейтральных аллелей даже в «бесконечно» большой популяции может выглядеть, как совершенно случайный процесс, хотя и приводимый в движение отбором.
Более того, процесс этот во многих отношениях аналогичен обычному дрейфу генов (конечно не по внутреннему механизму, а по внешнему проявлению) – те же блуждания частот аллелей до их окончательного закрепления в популяции. Отличие в том, что «селекционный дрейф» генов идет в любых по численности популяциях и под действием отбора, тогда как обычный дрейф – лишь в малых популяциях и «сам по себе». Естественно, что «селекционный дрейф» в зависимости от мощности отбора может охватывать одновременно множество генов, аллели которых в данный момент нейтральны друг относительно друга, и идет постоянно, никогда не прекращаясь, так как любые популяции постоянно находятся под действием отбора (во всяком случае, стабилизирующего – см. гл. 5).
Таким образом, нейтральная эволюция, в какой-то мере, может осуществляться не только за счет обычного дрейфа и мутаций (Кимура, 1985; Kimura, 1991), но и за счет своего рода «селекционного дрейфа» (попутного транспорта), подхватывающего мутации, удачно скомбинированные с «адаптивными» генами. Более того, можно предположить, что за счет попутного транспорта может в какой-то мере поддерживаться определенный уровень полиморфности популяций на подобие того, как это происходит за счет обычного дрейфа и мутационного давления в теории Кимуры (1985) только вне зависимости от численности популяций.2 Действительно, возникающие в популяции нейтральные мутации могут быть в ходе попутного транспорта либо элиминированы отбором, либо, наоборот, «подброшены» им до уровня полиморфных частот. Установившееся равновесие между этими противоположно направленными процессами и будет характеризоваться определенным уровнем полиморфности. Конечно, надо учитывать, что в действительности все это происходит на фоне действия также и обычного дрейфа.
При реальной эффективной численности популяции (Ne) скорее всего вклад последнего будет много более значительным чем рассматриваемого механизма. Но это всего лишь предположения. Как на самом деле может обстоять дело, и насколько значим попутный транспорт (и значим ли он вообще) для генетического преобразования популяций, ответить могут только математики, занимающиеся моделированием подобных ситуаций.
1 Процесс кроссинговера на самом деле не является случайным. Однако в масштабах тех участков хромосом, в которых эффективен попутный транспорт, его можно все-таки считать таковым.
2Подробно о механизмах поддержания полиморфизма см. в части 3.
113

ГЛАВА 8. СИСТЕМА СКРЕщИВАНИЯ
Микроэволюционные процессы в существенной мере зависят от «бытующей» в популяции системы скрещивания.
В простейшем случае скрещивание случайно (панмиксия). В большой популяции при этих условиях и при отсутствии действия отбора частоты аллелей неизменны, частоты генотипов находятся в равновесии, не изменяясь в последовательных поколениях (закон Харди – Вайнберга). Столь же неизменными остаются и распределения количественных признаков в популяции, если конечно среда, вызывающая модификационные изменения, не меняется. Действие на эту систему отбора изменяет частоты генотипов, но после первого же скрещивания (случайного) они опять приходят в равновесные состояния.
Однако популяции со случайным скрещиванием – абстракция. В действительности почти всегда (а может быть и всегда) скрещивание имеет неслучайный характер. Различают две основные формы неслучайного скрещивания: ассортативное скрещивание и скрещивание по родству. Ассортативное скрещивание может быть положительным, когда скрещивание особей с одинаковым фенотипом (генотипом по тому или иному локусу) происходит чаще, чем при случайном подборе пар, и отрицательным, когда скрещивание с одинаковым фенотипом (генотипом) происходит реже, чем при случайном подборе.
Вообще же любую популяцию со случайным скрещиванием можно разбить на две составляющие её группы. Одна образована парами особей с положительным ассортативным скрещиванием, другая − с отрицательным. Эту особенность иллюстрирует таблица 4.
Она отражает «процесс создания пар» при диаллельном скрещивании.
По диагонали таблицы затемнены пары с идентичными генотипами. Боковые элементы таблицы заняты парами с различающимися генотипами. Если образуются преимущественно «диагональные» пары, наблюдается положительное ассортативное скрещивание. Если «боковые» – то отрицательное скрещивание. Что же при этом может происходить на фено- и генотипическом уровнях в популяциях?
При положительном ассортативном скрещивании, когда «работают» только диагональные пары, частоты (p) аллелей равны (pА = pа = 0,5), генотипы в родительском поколении (Р) сформированы в ходе случайных скрещиваний (т.е. по Харди-Вайнбер- гу: pАА = 0,25; pАа = 0,5; pаа = 0,25), наследование кодоминантно (т.е. фенотипически гетерозиготы Аа отличимы от гомозигот АА) и поколения неперекрывающиеся, то последовательно (из «поколения» в «поколение» – F1, F2, …F∞) подставляя в таблицу 4 значения частот, получим следующий результат (табл. 5).
Таблица 4
Результаты случайного комбинирования самцов и самок в пары при диаллельном скрещивании
♀ |
♂ |
АА |
Аа |
аа |
|
||||
|
|
|
|
|
|
|
|
|
|
АА |
|
АА |
Аа |
аа |
|
|
АА |
АА |
АА |
Аа |
|
АА |
Аа |
аа |
|
|
Аа |
Аа |
Аа |
|
|
|
|
|
аа |
|
АА |
Аа |
аа |
|
|
аа |
аа |
аа |
|
|
|
|
|
114

|
|
|
|
|
|
Таблица 5 |
|
|
|
Изменение частот генотипов в гипотетической популяции |
|||||
|
|
|
при положительном ассортативном скрещивании |
||||
|
|
|
|
|
|
|
|
Генотип |
Р |
F1 |
|
F2 |
F3 |
F∞ |
|
|
|
|
|
|
|
|
|
АА |
0,25 |
0,3333 |
|
0,4167 |
0,4815 |
0,5 |
|
Аа |
0,5 |
0,3333 |
|
0,1667 |
0,0370 |
0 |
|
аа |
0,25 |
0,3333 |
|
0,4167 |
0,4815 |
0,5 |
|
Концентрация гетерозигот резко снижается уже к третьему поколению (F3), а в дальнейших поколениях асимптотически приближается к нулю. В конце концов, в популяции остаются только гомозиготы (pАА = pаа = 0,5). Частоты аллелей не изменяются (Р, F1, F2, … F∞ ; pА= pа = 0,5). Однако при таком типе скрещивания это состояние неустойчиво. Достаточно незначительного отклонения от равенства аллельных частот (pА≠ pа), чтобы в последовательных поколениях частота менее распространённого аллеля начала уменьшаться.
В конечном итоге в популяции зафиксируется наиболее распространённый аллель. При отрицательном ассортативном скрещивании, когда «работают» боковые элементы таблицы 4, а диагональные «молчат», в последовательных поколениях будет
наблюдаться совершенно иная картина.
При равенстве частот аллелей (pА = pа = 0,5) и исходном случайном скрещивании в родительском поколении (Р) частоты гетерозигот увеличатся в F1 до 0,6, а затем, испытывая небольшие колебания (от поколения к поколению), застабилизируются (практически уже через 3–4 поколения) на уровне 0,57.
При неравенстве частот аллелей (pА ≠ p а) будет также наблюдаться нечто подобное ― уровень гетерозиготности несколько возрастёт и, колеблясь, застабилизируется. Однако в данном случае, в отличие от предыдущего, не только частоты генотипов, но и частоты аллелей начнут изменяться (будет наблюдаться небольшое сопровождаемое колебаниями возрастание частоты редкого аллеля).
Врассмотренной выше модели предполагалось, что поиск партнёров осуществляется совершенно случайно, но скрещивания происходят только в парах, состоящих из адекватных партнёров. Неадекватные партнёры не скрещиваются, после чего дальнейший поиск партнёров прекращается. Это достаточно умозрительное предположение, но оно вполне может реализоваться в тех случаях, когда за период поиска возможно не более одной встречи между партнёрами (например, вследствие ограниченного времени поиска или малой численности особей). По-видимому более реально, когда партнёры неограниченно продолжают поиск и, в конце концов, по прошествии того или иного количества встреч обязательно находят нужного партнёра для скрещивания.1
Тогда при полном положительном ассортативном скрещивании частоты аллелей (какие быонинебылипервоначально)изменятьсянебудут,втовремякакгетерозиготностьбудет каждый раз (при переходе к новому поколению) двукратно уменьшаться. Если предположить, что гетерозиготы неотличимы по фенотипу от гомозиготАА (аллель А доминантен), принципиальных изменений не произойдёт. Частоты аллелей также не будут изменяться. Только уровень гетерозиготности будет уменьшаться медленней, чем в «кодоминантном» случае, так как часть рецессивных аллелей будет «укрываться» в гетерозиготах.
Вреальных условиях объединение пар и скрещивания могут протекать неким промежуточным способом между моделями с упорным поиском партнёров (т.е. с множеством их встреч) и без него (т.е. с не более чем одной встречей партнёров).
1 Во всяком случае именно эта модель обычно рассматривается в учебниках по популяционной генетике (см. например, Ли, 1978; Хедрик, 2003).
115

Более того ассортативность может быть далеко неполной, а особи могут лишь слегка предпочитать (или не предпочитать) тот или иной фенотип. То есть возможно существование великого множества вариантов, как положительного, так и отрицательного ассортативного скрещивания, изменяющих и не изменяющих аллельных частот и приводящих к различным изменениям частот генотипов по одному, двум и большему числу локусов.
Естественно, что ассортативность может проявлять себя не только по качественным признакам (рассмотренным выше), но и по количественным (точнее параметрическим) как с плавным, так и дискретным проявлением. Типичное их проявление наблюдается при положительном ассортативном скрещивании (вспомним подбор пар у человека, который осуществляется отчасти именно таким способом − Эрман, Парсонс, 1984). С помощью всё той же «диагональной» модели (табл. 4, 5) можно очень грубо проиллюстрировать и этот случай.
Примем, что три генотипа аа, аА, АА контролируют, например, такие значения некоего гипотетического признака – 0; 0,5; 1. Тогда после начала ассортативного скрещивания обнаружится, что у нашего признака, при неизменности среднего его значения, начнёт быстро нарастать изменчивость, выраженная через среднеквадратическое отклонение от 0,35 (в родительском поколении – Р) до предельного значения 0,5 (достигаемого практически уже через 3–4 поколения). Коэффициенты корреляции при этом между скрещивающимися фенотипами равны единице 2.
Более реальная модель должна учитывать наличие модификационной изменчивости, которая переводит признак с дискретными значениями (0; 0,5; 1) в состояние с плавным варьированием. В силу только этого, «точность» ассортативного объединения генотипов понижается (т.е., уровень ассортативности скрещиваний падает), вследствие чего резко понижается значение коэффициента корреляции, с помощью которого и измеряется уровень ассортативности. Изменчивость при этом нарастает не столь интенсивно. Наконец, чтобы ещё более приблизиться к реальности, желательно перейти к модели с «упорным поиском партнёров», увеличить число аллелей и локусов, а также ввести некоторые другие дополнения.
При отрицательных скрещиваниях изменчивость количественного признака будет уменьшаться, а среднее его значение может не только оставаться неизменным, но и увеличиваться или уменьшаться (всё это легко представить себе, исходя из той же «диагональной» модели).
Рассмотренными выше простейшими вариантами положительного и отрицательного ассортативного скрещивания, конечно же, не исчерпываются все возможности выборочного объединения пар. Мыслимы и многие другие варианты. Например, преимущественное скрещивание самцов и/или самок с гетерозиготами; преимущественное скрещивание гетерозигот; предпочтительное скрещивание с одной из гомозигот.
Впервом и втором случаях в последовательных поколениях будет нарастать гетерозиготность (до определённого уровня), а частоты аллелей будут стремиться к равновесным значениям (равным 0,5).
Впервом случае всё это будет происходить медленнее, во втором – быстрее. В третьем случае, напротив, будет наблюдаться снижение уровня гетерозиготности в попу-
2 з аметим: реально обнаруживаемые коэффициенты корреляции обычно ниже 0,5, что вполне понятно, так как на самом деле популяция может состоять из особей с разными «вкусами», т.е. из предпочитающих либо случайные скрещивания, либо положительные, либо отрицательные. Комбинация этих «вкусов» и создаёт общую картину. Кроме того, «вкусы» особей могут изменяться в процессе их жизни, приводя к зависимости коэффициента корреляции от возрастного состава. Немаловажное значение имеет и способность особей дифференцировать фенотипы. Ещё одна причина – модификационная изменчивость (см. последующий текст).
116

ляции и увеличение частоты того аллеля, из которого составлены предпочитаемые гомозиготы.
Еслипредпочтениябудутвсякийразисчезатьвпроцессевозрастаниячастотыаллеля или меняться в пользу альтернативных гомозигот, возникнет состояние с устойчивым равновесием частот аллелей в популяции. Казалось бы все эти гипотетические случаи (приводимые здесь только для иллюстрации возможностей ассортативного скрещивания) малореалистичны. Однако в той или иной мере иногда они, по-видимому, всётаки могут реализоваться. Например, ещё с 60-х годов известны экспериментальные подтверждения возможности преимущественного скрещивания самок дрозофилы с самцами редкого типа. Когда такой тип самцов становится распространённым, преимущества исчезают. Предполагается, что подобный механизм может действовать не только у ряда видов насекомых, но и в отдельных случаях у некоторых видов птиц и других представителей животного мира.
Как можно было заметить, ассортативное скрещивание в различных своих вариантах является по существу половым отбором3. К сходным последствиям оно может приводить и при ассортативности объединения гамет.
Эффекты, возникающие при скрещивании по родству, также можно пояснить на простой модели (табл. 6).
здесь изображен наиболее общий случай скрещиваний между «двуполыми», т.е. гермафродитными (♂♀) особями (а1, a2, a3, …aN). На диагонали, выделенной тоном, лежат комбинации скрещивания особей «самих с собой» (самооплодотворение). Вправо от диагонали указаны комбинации, когда первая особь выступает как самец. Влево − когда первая особь выступает как самка. Кроме того, полагается, что все особи в таблице 6 расположены не хаотично, а по родству, которое убывает по мере удаления их друг от друга.
Таблица дает общую картину всего многообразия скрещиваний в популяции при их случайном комбинировании. Но эти случайные комбинации можно разделить на три категории: скрещивания с самооплодотворением, скрещивания родственных особей (ближайшие номера пар в таблице), скрещивания неродственных особей (соответственно, дальние номера пар).
Если вдруг такая популяция перейдет от случайного скрещивания исключительно к самооплодотворению (зачеркиваем все ячейки в таблице 6, кроме диагональных), то постепенно от поколения к поколению доля гетерозигот (Н) в популяции будет неуклонно, как Hn = (1/2)n H0 (H0 − исходная гетерозиготность, Hn – гетерозиготность в n−ом поколении), стремиться к нулевому значению, генетическая дисперсия (как, впрочем, и общая фенотипическая изменчивость) будет нарастать, а частоты генотипов в популяции будут неуклонно приближаться к частотам генов и в пределе (n→∞) сравняются с ними.
|
|
|
|
Таблица 6 |
|
|
|
Комбинации скрещивающихся условных особей |
|||
|
|
|
|
|
|
♂♀ |
а1 |
a2 |
a3 |
…aN |
|
♀♂ |
|
|
|
|
|
а1 |
а1 а1 |
a2 а1 |
a3 а1 |
… а1 aN |
|
a2 |
а1 a2 |
a2 a2 |
a3 a2 |
… a2 aN |
|
a3 |
а1 a3 |
a2 a3 |
a3 a3 |
… a3 aN |
|
… aN |
… aN а1 |
… aN a2 |
… aN a3 |
… aN aN |
|
3 Предыдущий вариант скрещивания самок с самцами редкого фенотипа представляет собой частотно-зависимый половой отбор.
117
Когда популяция представлена не практикующими самооплодотворение однополыми особями, из таблицы 6 необходимо вычеркнуть диагональные элементы и рассматривать уже только две оставшиеся категории. В этом случае для любого локуса можем записать так,какэтопредставленовтаблице7,вкоторойчерезp, q обозначенычастотыA,ааллелей, а через F− коэффициент инбридинга, изменяющийся от 0 до 1 и характеризующий свойства «реальной» популяции (т.е. популяции с инбридингом − скрещиванием родственных особей) в сравнении со свойствами «идеальной» (бесконечно большой) панмиктической популяции, в которой встреча родственников бесконечно мала (для нее частоты генотипов: p2,2pq, q2−распределение Харди−Вайнберга). По существу также можем поступить, когда популяция представлена не гермафродитными, а «нормальными» особями. Для этого надо из таблицы 6 вычеркнуть дополнительно и все наддиагональные элементы. Тогда останутся лишь самки (первые элементы в таблице), скрещивающиеся с самцами (вторые элементы в таблице). Естественно можно поступить и наоборот. Так как ситуация симметрична, получим один и тот же результат, в любом случае включая и «гермафродитизм».
Если мы вычеркнем из модифицированной таблицы 6 (с однополыми особями) все неродственные друг другу пары, т.е. оставим лишь скрещивания между родственниками, которые лежат на диагоналях, примыкающих к изъятой только что диагонали (с самооплодотворением), то получим усиление инбридинга в популяции (значительное увеличение F и, соответственно, снижение уровня гетерозиготности). При продолжительном существовании такой популяции (в которой скрещивания идут всякий раз между родственниками) события буду развиваться примерно так, как это было описано выше для популяции с самооплодотворением (но помедленнее).
Если же мы вычеркнем из той же таблицы 6 всех родственных особей, оставив категорию неродственных, F превратится в нуль и, соответственно, гетерозиготность возрастет до величины, характерной для панмиктической популяции (с поправкой на случайные процессы).
Обе эти крайние ситуации (родственных и неродственных скрещиваний) могут быть встречены в природе, но наиболее характерны для популяций, контролируемых человеком, где инбридинг и аутбридинг являются одними из форм разведения, применяемых в селекции.
заметим, что в отличие от ассортативного скрещивания, скрещивание по родству определяет поведение всей совокупности генов организма, а не отдельных их групп или единиц.
С. Райт (Wright, 1951) заметил, что преобразованное с учётом родственных скрещиваний распределение Харди-Вайнберга (табл. 7) по форме совпадает с тем же распределением но полученным для популяции, подразделённой на субпопуляции (табл. 8).
В этой таблице через V обозначена дисперсия частот генотипов. Данное распределение (табл. 8) характеризует так называемый эффект Валунда (Wahlund, 1928) – снижение частоты гетерозигот от «стандартно» ожидаемой величины (по Харди-Вайнбергу) при переходе популяции к подразделённому «образу жизни».
Далее нетрудно было заметить, что не только по форме, но и по содержанию формулы в обеих таблицах по существу отражают почти одно и то же. Действительно, с
|
|
|
Таблица 7 |
|
|
|
Генотипы и их частоты в «реальной» популяции |
||
|
|
|
|
|
|
|
Уровень |
|
|
|
|
гетерозиготности (Н) |
|
|
Генотипы |
AA |
Aa |
аa |
|
Их частоты в популяции |
p2+Fpq |
2pq – 2pqF |
q2+Fpq, |
|
118

Таблица 8
|
Частоты генотипов в популяции, подразделённой на субпопуляции |
|||
|
|
|
|
|
Генотипы |
AA |
Aa |
aa |
|
Частоты |
p2+V |
2pq-2V |
q2+V |
|
одной стороны, подразделение большой популяции на множество мелких приводит к увеличению числа скрещиваний генетически близких особей. С другой – состояние не подразделённой с виду популяции с идущими в ней родственными скрещиваниями может быть интерпретировано как своеобразное подразделение (изоляция расстоянием). Отсюда остаётся только приравнять формулы из обеих таблиц друг другу и получить окончательный результат:
Таким образом, оказалось, что с помощью коэффициента инбридинга вполне можно описывать генетическую дифференциацию популяции, действительно разбитой на субпопуляционные группировки. Разрабатывая эту теорию, С. Райт ввёл несколько типов коэффициентов инбридинга, по-разному характеризующих генетическую структуру популяции.
FIT – коэффициент инбридинга особи (I) в целой популяции (Т);
FIS – коэффициент инбридинга особи (I) в субпопуляции (S);
FST – коэффициент инбридинга субпопуляции (S) в целой популяции (T). Коэффициенты связаны соотношением
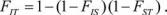 .
.
Каждый из них по-своему отражает отклонение от панмиксии, возникающее либо из-за подразделённости популяции (FST ), либо из-за идущих в субпопуляциях близкородственных скрещиваний (FIS )4. Совокупное действие обоих эффектов передаётся FIT-коэффициентом.
Подробности и способы расчёта коэффициентов5 можно найти почти в любом руководстве по популяционной генетике.
4Или из-за их подразделённости.
5 Для расчётов необходимы лишь сведения по частотам аллелей и генотипов в популяциях и субпопуляциях.
119