

Рожков Ю. И. Популяции, виды, эволюция
.pdfОднако всё это ещё не самое «страшное» в физике хуже всего то, что две базовые для этой науки теории (теория относительности и квантовая механика) строятся на активном участии наблюдателя с его измерительными приборами в познании физической картины мира. Причём, если в классической физике человек - это сторонний наблюдатель, изучающий мир, существующий сам по себе, то в «новой» физике этот мир (или микромир) приобретает всякий раз несколько иное обличие в зависимости от поведения наблюдателя (отсылаем читателей за подробностями к специальной литературе).
Эти странности взаимодействия «физики» и «разума» настораживают, и похоже, что с годами развития науки подобные взаимодействия всё более будут крепнуть3.
Не свидетельствует ли это о том, что мы находимся где-то у начала процесса, который, уходя в бесконечность, откроет неразделимое слияние того и другого? Конечно же, это чистейшая полурелигиозная фантазия, возможно достойная всякого порицания.
Однако допустим на минуту, что это действительно так и есть. А те «несуразности», которые нам демонстрирует сегодня точнейшая из современных физических наук - квантовая механика, это лишь первые предвестники такого слияния. Тогда придётся допустить, что в далёком будущем биологи, занимающиеся фундаментальными проблемами (в том числе и проблемами разума), не исчезнут, а будут всегда находиться в поиске за пределами круга4. Эволюционисты же откроют какие-то совершенно небывалые эволюционные факторы развития органического мира, позволяющие легко объяснить то, что очень трудно объяснить сейчас.
Нам же остаётся, заканчивая фундаментальный раздел книги (части 1-4), приступить в следующем, последнем разделе (часть 5) к прикладным и полуприкладным проблемам, то есть углубиться внутрь круга.
ЧАСТЬ V. ЭВОЛЮЦИЯ, ИНИЦИИРУЕМАЯ ЧЕЛОВЕКОМ
Содной стороны человек, исходя из своих потребностей, активно преобразует и создаёт породы животных, сорта растений и различные варианты микроорганизмов.
Это, так сказать, его осознанная деятельность по проведению искусственной эволюции с целью получения форм способных наиболее эффективно его накормить, приодеть, подлечить и даже развлечь.
Сдругой стороны он борется "за урожай" с различными "нахлебниками (насекомыми, грызунами, сорными растениями, микроорганизмами и т.д.) мешающими ему присвоить исключительно себе всё вышеперечисленное. В этой борьбе он использует инсектициды, гербициды, различные яды для млекопитающих, антибиотики для патогенных бактерий и прочее. Тем самым "неосознанно" инициируя эволюционный процесс в противодействующих ему популяциях. Изучением подобных явлений активно занимались ещё в середине прошлого века (в 60-х годах начали выходить даже монографии на эту тему).
В80-х было подсчитано, что в мире (на то время) существовало более 500 различных видов насекомых и клещей вредителей сельскохозяйственных культур ставших устойчивыми к пестицидам в результате их отбора на приспособленность к применяемой "отраве" (Яблоков, Остроумов, 1985). Исходя из общей тенденции увеличения со временем число приспособившихся видов, эта цифра должна быть гораздо выше. Если же подсчитать не только вредителей, но и тех которые просто страдают с ними вместе "за компанию", цифра должна возрасти гигантски.
Всё это и многое другое очень хорошо освещено в научно и учебной литературе. Кое какие из подобных фактов приводили и мы в части III нашей книги. Поэтому отсылаем читателей к многочисленным трудам подобного рода.
Мы же здесь сосредоточимся совсем на другом, на том что гораздо менее известно и не столь широко дискутируется научной общественностью – на "манипуляциях" человека действующего "бессознательно", как истинный селекционер в природных популяциях и сознательно в популяциях сельскохозяйственных и домашних животных.
3То есть, новые базовые физические теории будут «страннее» предыдущих.
4Кстати, из всего этого следует, что разум столь же непознаваем (или вернее - бесконечно познаваем) как и Вселенная (т.е. физика). Возможно это обрадует религиозно настроенных читателей.
300
ГЛАВА 22. ДЕЙСТВИЕ ПРОМЫСЛОВОГО ОТБОРА В ПОПУЛЯЦИЯХ
Одним из актуальных приложений теории эволюции, возникших сравнительно недавно, является оценка роли промыслового воздействия на популяции (Рожков, Проняев, 1994). Прямое преследование (в том числе и охота) как фактор деградации видов последние десятилетия отошло на второй план, пропустив вперёд такую сборную группу факторов, как трансформация местообитаний (World Cons. Strategy, 1980; Конвенция 1995). Однако виды, сохраняющие текущую экономическую ценность, испытывают двойное воздействие. С одной стороны, из их популяций ежегодно изымается сравнимое по численности с приростом поголовье, с другой - их местообитания изменяются, а в худших ситуациях - уничтожаются со скоростью, превышающей адаптивные способности вида. Хотя фактор промысла/охоты в среднем стал менее значимым в процессе деградации видов, понимание механизма его действия крайне важно, так как может способствовать смягчению воздействия или, в ситуациях с некоторыми видами, даже снятию его влияния.
На смежении 80-х и 90-х годов прошлого столетия мы обратили внимание на характер промыслового воздействия в отношении внутрипопуляционной изменчивости (Проняев, Рожков, 1988; Проняев, Аксёнов, 1988; 1991; Рожков, Проняев, 1994; Рожков и др., 2001). Следуя традициям российской научной школы, мы предложили механизм этого воздействия (на основе введенного принципа эффективности), позволяющего утверждать, что практически любое промысловое изъятие из популяций носит направленный характер (Рожков, Проняев, 1994). В последнее время и в англоязычной научной литературе появились работы, подтверждающие направленный характер изъятия, существенно ускоряющий фенотипическое преобразование популяций, используемых человеком в качестве источников белка, сырья или развлечения
(Hendry et al., 2008; Darimont et al., 2009). Однако в публикациях зарубежных коллег нами не встречено объяснительного начала направленности воздействия, за исключением трофейных охот. В основном обсуждаются результаты этого процесса: скорость преобразования признаков и возможные последствия. Поэтому мы сочли необходимым сохранить и воспроизвести логику изложения нашей публикации 1994 года, позволяющей уяснить формирование механизма возникновения бессознательного отбора при использовании свободноживущих популяций, дополнив изложение некоторыми новыми фактами, полученными непосредственно нами или почерпнутыми из литературы.
22.1. Элементарное введение для исследователей, далёких от эволюционных проблем
Естественный отбор признается большинством исследователей как ведущий эволюционный фактор, с одной стороны, поддерживающий видовые особенности, с другой - формирующий на основе генетически обусловленной изменчивости новые признаки и свойства популяций, что, в конечном итоге, может завершаться образованием и новых видов. Действие отбора заключается в двуедином процессе дифференциальной выживаемости и дифференциальной плодовитости особей вида. Сохранение особи, не способной размножаться или дающей меньше потомков, чем средняя особь в популяции, не приведёт к увеличению числа наследственных задатков этой особи в последующих поколениях (подробнее см. гл. 5).
В современных популяциях эксплуатируемых видов одним из ведущих в численном отношении факторов элиминации животных выступает промысел/охота. По усредненным оценкам, ежегодноизпопуляций этихвидовизымаетсясравнимое сгодовымпри-
301
ростом, а в некоторых ситуациях – и большее, число животных. Очевидно, что столь мощный элиминирующий фактор должен привлечь внимание хотя бы по той причине, что оказывает прямое воздействие на выживаемость особей в популяции и опосредованно может влиять на дифференциальную плодовитость посредством сохранения и/ или уничтожения в популяции тех или иных генотипов, то есть влиять на биологическое разнообразие вида. Однако в большинстве случаев в настоящее время за редким исключением контроль изъятия проводится лишь с позиций численности и/или плотности опромышляемого вида (Давыдов, Рожков, 2009)1.
По всей видимости, такой подход является отражением исторического пути становления научных основ рыболовства, охотоведения (game management), когда ресурсы того или иного вида рассматривались с позиций экологии, точнее популяционной экологии. При этом во внимание принимались и принимаются до сих пор статические и динамические параметры популяций, не учитывающие разнокачественность особей. Собственно, это основа популяционно-экологического мышления (Гиляров, 1991).
Развитию и становлению такого подхода способствовала и антропоцентрическая система взглядов на природу в целом и отдельные биологические виды в противовес развивающемуся биоцентрическому подходу в рамках современной биоэтики. Антропоцентрическая система взглядов рассматривает ресурсы вида прежде всего как источник белка или иного сырья (удовольствия, отдыха), а не как уникальную эволюционно сложившуюся биологическую систему, входящую в глобальную экологическую систему – Биосферу. зачастую собственно виды как элементы глобальной системы более ценны, чем продукция, получаемая от них в настоящее время (Конвенция…, 1995).
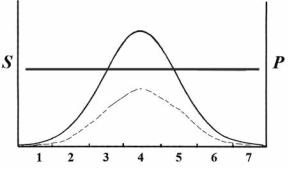
В господствующем в настоящее время практическом подходе априорно принимается, что все изымаемые особи одного возраста и пола идентичны по своим фено- и генотипическим характеристикам (что абсолютно неверно), или что изъятие не зависит от тех или иных качеств особи (не избирательно2). Последнее высказывание может быть проиллюстрировано (рис. 123).
Рис. 123. Господствующее представление о безвыборочном характере промысловой элиминации. жирной линией обозначена кривая элиминации; тонкой – распределение признака в популяции до элиминации; прерывистой – после элиминации. S – коэффициент элиминации; p
– частота; 1, 2, … 7 – классы признака.
|
|
|
1 |
В ряде схем эксплуатации предусматривается оценка и регуляция возрастного и полового |
|
состава изымаемых особей. |
||
2 |
Очевидно, что при трофейных охотах учитывается разнокачественность особей, по крайней |
|
мере, в отношении тех признаков, которые характеризуют трофей.
302

Из популяционной биологии (морфологии, генетики) известно, что внутри каждого возрастно-полового класса существует разнообразие (изменчивость) как по количественным, так и по качественным признакам. На рисунке 123 изображено частотное распределение некоего количественного признака одного возрастно-полового класса в популяции. В большинстве известных случаев это распределение близко к нормальному, поэтому оно и отражено в виде колоколообразной кривой. Прямая, паралелльная оси абсцисс – коэффициент промысловой элиминации. Следует дать пояснение, что понимается под элиминационной кривой.
Для представления изменчивости непрерывно варьирующих (количественных) признаков используют разбиение всего массива данных по тому или иному признаку на классы. Собственно колоколообразная кривая и построена на основе частот встречаемости особей в каждом из выделенных классов, вычисленных как количество особей, приходящихся на тот или иной интервал, отнесенное ко всему объему
данных ( nNi , где ni – количество особей в i-том классе, N – объём выборки).
Коэффициент элиминации определяется как отношение числа выбывших особей в классе к числу особей в нём до выбытия (отбора) (подробней об этом см. Приложение 4). Рассмотрим пример расчета коэффициента элиминации (табл. 17).
Как следует из рисунка 123 и таблицы 17, в любой зоне распределения признака изъятие равновероятно. Исходя из этой модели, после промысла распределение признака не трансформируется, а происходит пропорциональное сокращение численностей особей во всех зонах распределения (рис. 123).
Очевидно, что при таком понимании воздействия охоты (промысла) обсуждать какие-либо эффекты на уровне популяций в рамках микроэволюционных воззрений было бессмысленно. По всей видимости, именно господство этой модели в умах исследователей, далёких от эволюционных проблем, и тем более практиков не приводило к изучению процессов, связанных с промысловой элиминацией. Для нашей страны сохранение подобных взглядов видимо могло сформироваться из-за известных событий в биологии, когда отрицалось само представление об изменчивости и наследственности и их роли в эволюционных процессах (Медведев, 1992).
Вместе с тем, модель (рис. 123; табл. 17) безвыборочности элиминации представляет собой один из бесконечного числа вариантов расположения элиминационных кривых (рис. 124). Предпочтение модели, отображённой на рисунке 123, или иных моделей, отображённых на рисунке 124, не имеет каких-либо научных оснований.
Вероятно, приведенные выше представления о характере промысловой элиминации не привлекли бы внимания, если бы в разных систематических группах эксплуатируемых видов не наблюдались сходные процессы, а именно: измельчание, сдвиг полового созревания, сокращение плодовитости или ее резкий рост, увеличение ранней постнатальной смертности молодняка, изменение уровня фенотипической и генетической изменчивости, изменение сроков миграционной активности, изменение сроков появления молодняка и некоторые другие.
Таблица 17
Пример расчёта коэффициента элиминации для рисунка 123
Кол-во |
|
|
Классы признака |
|
|
Сумма |
|||
|
|
|
|
|
|
|
|||
животных, особей |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
||
|
|||||||||
|
|
|
|
|
|
|
|
|
|
Элиминировано |
3 |
5 |
10 |
15 |
10 |
5 |
3 |
51 |
|
До элиминации |
6 |
10 |
20 |
30 |
20 |
10 |
6 |
102 |
|
Коэф. элиминации |
0,5 |
0,5 |
0,5 |
0,5 |
0,5 |
0,5 |
0,5 |
|
|
303

Рис. 124. Множество теоретически возможных ориентаций элиминационных кривых относительно распределения признака. Условные обозначения те же, что и на рисунке 123.
Примечателен тот факт, что изменение параметров обнаруживается в разных систематических группах животных, обитающих в разных физических средах, имеющих разные механизмы взаимодействия со средой, но объединенных одним фактором – промысловым использованием. Чтобы не быть голословными, в следующем разделе приведем примеры ряда отслеженных ситуаций послепромыслового изменения параметров популяций.
22.2. Примеры, свидетельствующие о направленном характере промысловой элиминации
По вполне понятным причинам, для промысловых популяций, в отличие от сельскохозяйственных или лабораторных, трудно получить прямые результаты, которые свидетельствовали бы о характере направленности промысловой элиминации. Это связано с тем, что в контролируемых группировках животных можно проследить судьбу особи (группы) от рождения до выхода из популяции. В свободноживущих популяциях возможно выявить только общие тренды поведения весо-ростовых признаков при налаженном мониторинге3.
Большинство приводимых примеров иллюстрируют конечные следствия воздействия промысла. При этом следует иметь в виду, что отслеживаемые признаки могут и не быть непосредственно теми признаками, по которым происходит изъятие особей. Скорее всего, они связаны коррелятивно с особенностями биологии их носителей.
Ниже мы преднамеренно стремились привести примеры, с одной стороны, касающиеся видов разных систематических групп животных, а с другой – видов, имеющих разные стратегии взаимодействия со средой, с тем, чтобы убедить читателя в объективности существования направленной промысловой элиминации. Во всех примерах мы не будем останавливаться на деталях, которые достаточно полно описаны в первоисточниках.
Первая группа примеров относится к эксплуатации ресурсов популяций рыб. Примеры для этой группы наиболее представительны (Алтухов, 1989, 2003; Алтухов и др., 2004; Матишев, 1990а, б; Ricker, 1981; Vaughan, 1947 и многие др.). Это связано с тем,
3 Бесспорно, для наземных свободноживущих позвоночных можно предложить крайне дорогостоящий (а потому и неосуществимый) проект с массовым мечением спутниковыми передатчиками и отслеживанием индивидуальной судьбы особей с известными фено-, генотипическими параметрами от рождения вплоть до выбытия из популяции.
304
что в питании человека 20% потребляемых белков рыбного происхождения. Среднестатистический житель земли потребляет около 19 кг рыбы ежегодно. запасы рыбы интенсивно, в промышленных масштабах эксплуатируются более ста лет и для многих стран являются основой экономического процветания. Ежегодные мировые уловы оцениваются в 100 млн. т (Голубев, 1999). Поэтому к оценке состояния рыбных запасов привлечено большое количество исследователей.
Рассмотрим детально проанализированную Ю.П. Алтуховым и его сотрудниками (Алтухов, 1989; 2003; Алтухов и др., 2004) популяцию нерки (Ocorhynchus nerka) оз. ДальнегонаКамчатке,служащегообъектомдлительногомониторинга(Крогиус,1979).
Вследствие промышленного лова изменились следующие параметры популяции: сократилась численность заходящих в озеро производителей с 62.6 тысяч экземпляров (1935–46 гг.) до 1.6 тысяч (1966–76 гг.); произошло увеличение доли мелких самцов («каюрок») среди проходных самцов с 0.2% (1935–46 гг.) до 37.5% (1966–76 гг.); увеличилась доля карликов среди всех самцов с 26% (1935–46 гг.) до 88.8% (1966–76 гг.); возросла доля самок в проходной части стада с 52% (1935–46 гг.) до 68% (1966–76 гг.).
Указанные изменения сопровождались также трансформацией эволюционно сложившейся генетической структуры популяций из-за интенсивной промысловой элиминации более гомозиготных особей.
Во многом сходные изменения отмечены для популяций нерки озёр Ближнего и Начикинского (Алтухов, 1989; 2003; Алтухов др., 2004). Такую реакцию популяций лососей авторы связывают с интенсивным промыслом, в первую очередь, раннемигрирующих субпопуляций, в которых сконцентрированы наиболее крупные, более гомозиготные рыбы, среди которых преобладают самцы.
Другой пример относится к популяциям горбуши (Ocorhynchus gorbuscha) на юговостоке Аляски (Vaughan, 1947, цит. по Майру, 1968). Многолетние наблюдения за датой, к которой вылавливалось 75% общего улова, выявили устойчивую тенденцию отодвигания этого события на более поздние сроки. Например, для одной из популяций 75% улова было добыто в 1921 г. 28 июля; в 1926 г. – 2 августа; в 1930 – 9 августа; в 1936 г. – 16 августа; в 1944 г. – 18 августа. Отсюда вывод – промысел приводит к практически полной элиминации особей с генами (аллелями), определяющими ранний нерест.
Исследования горбуши и чавычи, проведённые Риккером (Ricker, 1981) в водах Британской Колумбии, основанные на многолетнем мониторинге, однозначно свидетельствуют о направленной элиминации при промысле этих рыб (рис. 125, 126). Автору даже удалось вычислить коэффициенты наследуемости и селекционные дифференциалы при различных технологиях лова.
Следующий пример относится к популяциям трески и ряда других видов океанических рыб Баренцева моря (Матишев, 1990а, б). Результатом промышленного лова явилось значительное изменение возрастной структуры популяции в сторону ее омоложения. К 1970-му году в уловах перестали регистрироваться 15–20-летние особи, к 1980-му году фактически не осталось 10–15-летних рыб. Основу современной добычи составляют 3–6-летние рыбы. При этом возросли темпы полового созревания. В 50–60-ых годах половозрелость наступала у 8–10-летних особей, в настоящий момент обнаруживается 5–20% половозрелых рыб 3–5-летнего возраста. Раннее участие в размножении сопровождается сокращением плодовитости (количества выметываемой икры). Кроме того, обнаруживается и уменьшение линейных размеров трески.
Аналогичные по характеру изменения обнаруживаются и в популяциях сельди и мойвы. Для первой, кроме того, характерно изменение сезонной смены стаций. В прошлом нагул особей этого вида происходил у берегов Норвегии, в настоящее время этого не отмечается (Матишев, 1990б).
305
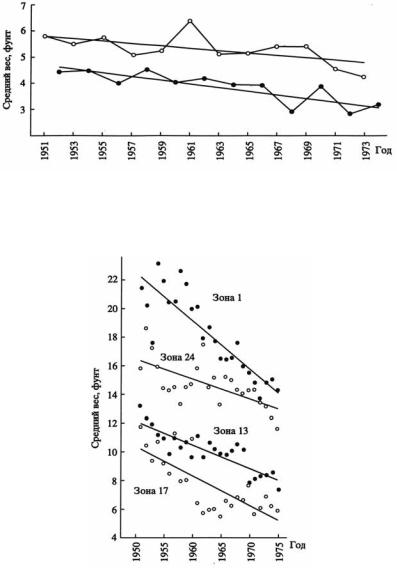
Рис. 125. Изменение среднего веса горбуши (по: Ricker, 1981 с изменениями). Поколения нечётных лет – светлые кружки; чётных – чёрные.
Рис. 126. Изменение среднего веса чавычи (по: Ricker, 1981).
Автор считает, что одной из основных причин сложившейся ситуации, ведущей к вырождению эксплуатируемых видов рыб, является промысел, который благоприятствует сохранению рыб меньшего размера, имеющих меньший темп роста, воспроизводства. Ряд ситуаций, касающихся селективного действия промысла рыб, проанализированы также и другими авторами (Law, 2000).
Во второй группе собраны примеры, касающиеся копытных животных. Эти животные испытывают огромное негативное влияние со стороны человека. В историческое время (сравнительно недавно) под воздействием человека исчезли тарпан, тур. Лошадь Пржевальского, зубр, бизон, олень Давида в настоящее время сохраняются в ограни-
306
ченных по численности группах (Банников и др., 1999; Немцев и др., 2003; Спасская, 2001). значителен список копытных, занесённых в Красную книгу. Такое состояние этой группы животных имеет несколько причин. С одной стороны, копытные как исторически сложившийся источник белка у многих народов (или развлекательной охоты) испытывают на себе воздействие интенсивного промысла. С другой – наблюдается существенное преобразование местообитаний этих животных или сокращение площадей, а во многих ситуациях и уничтожение среды их обитания. Традиционная мера
– создание особо охраняемых территорий, не всегда действенна для копытных. Для существования самовоспроизводящихся групп необходимы более значительные по площади пространства, нежели площади современных заповедников, национальных парков и заказников (Биология охраны…, 1983; жизнеспособность …, 1989).
Первый пример относится к изолированной (калмыцкой) популяции сайгака (Saiga tatarica), обитающей на правобережье Волги. С начала 80-х годов прошлого столетия добычу животных в ней стали осуществлять с применением переносного кораля
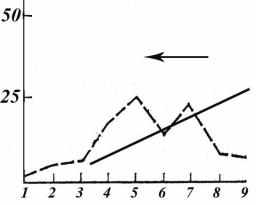
– высокоэффективной технологии, позволяющей за день промысла добывать несколько тысяч животных (Максимук, 1982). В условиях ограниченных по времени сроков промысла и стремления максимизировать доходы, добыча животных из мелких, спорадично расположенных стад (а тем более, одиночных животных) с применением кораля, как правило, не производилась, так как неэффективна. В терминах векторной теории отбора это может быть представлено следующим образом (рис. 127).
Из популяции, как правило, изымаются животные, образующие очень крупные, крупные и средние по размеру стада. В результате промыслового отбора в популяции сохраняются животные, ведущие одиночный или мелкогрупповой образ жизни.
В середине 80-х годов прошлого века при сравнительном анализе текущих параметров калмыцкой и казахстанской популяций, а также при их сопоставлении с данными по калмыцкой популяции конца 50-х годов было отмечено, что животные калмыцкой популяции стали иметь инфантильные черты как по количественным признакам эксте-
Рис. 127. Модель отбора, наводимого по признаку размер стад при применении сетевого кораля. По шкале ординат – доля стад различного размера. По оси абсцисс – размер стад: 1 – от 1
до 5 особей; 2 – 6–10; 3 – 11–19; 4 – 20–49; 5 – 50–100; 6 – 101–200; 7 – 200–500; 8 – 501–1000; 9 – более 1000 особей. Пунктирной линией обозначено частотное распределение размеров стад в сентябре по Л.В. жирнову (1998). Сплошной линией показана элиминационная кривая; стрелкой указано направление действия отбора.
307
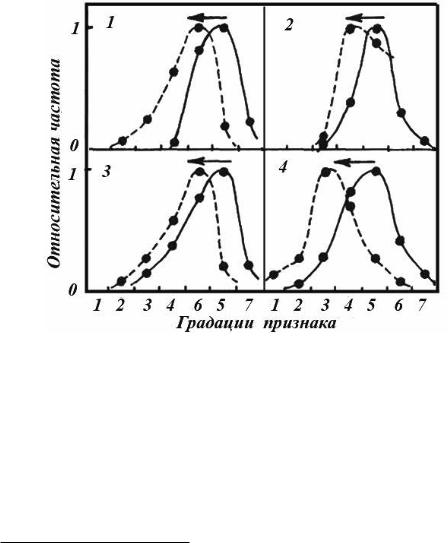
рьера, так и по характеристикам черепа (Проняев, 1985). Несколько позже было обнаружено резкое сокращение генотипического и фенотипического разнообразия (Проняев, Рожков, 1988; Проняев, Аксенов, 1988). В те же годы при детальном анализе экстерьера особей в последовательных промысловых сезонах выявлен направленный характер промыслового изъятия (Проняев, Аксенов, 1991). Из каждого возрастнополового класса животных в результате промысла изымались наиболее развитые особи (рис. 128). То есть промысел наводил во всех возрастно-половых группах минус-вектор отбора. Примечательно, что во всех четырех парах сравнений действие искусственного отбора инвариантно по направлению4.
Как следствие действия направленного отбора в популяции можно рассматривать значительное сокращение эмбриональной плодовитости у взрослых самок в результа-
Рис. 128. Действие промысловой элиминации в популяции сайгака. По горизонтали: в 1–3 градациях признака – косая длина туловища (см) у самок: 1 – 48–51; 2 – 52–55; 3 – 56–59; 4 – 60–63; 5 – 64–67; 6 – 68–71; 7 – 72–75; в 4 градации признака – обхват груди за лопатками (см) у поло-
возрелых самцов: 1 – 73–75; 2 – 76–78; 3 – 79–81; 4 – 82–84; 5 – 85–87; 6 – 88–90; 7 – 91–93. По вертикали: относительная частота встречаемости животных (отнормированная на максимальное значение). 1 – сплошной линией изображено распределение признака у взрослых животных (старше 2.5 года) в 1986 г., пунктиром – та же группа после промыслового воздействия; 2
–сплошной линией изображено распределение признака у 18-месячных животных, пунктиром
–та же группа через 6 месяцев; 3 – сплошной линией изображено распределение признака у взрослых животных после воздействия промысла в 1985 г., пунктиром – та же группа после промыслового воздействия 1986 г.; 4 – сплошной линией изображено распределение признака до промысла, пунктирной – та же группа после промысла. Стрелкой указано направление действия в популяции искусственного отбора.
4 Позже нами был проведен анализ отбора и по комплексу признаков («суперпризнак»), который показал аналогичные результаты (Проняев, Рожков, 1998) – гл. 5.
308

те практически полного отсутствия двойневых беременностей, столь характерных как для казахстанских популяций вида, так и для калмыцкой популяции конца 50-х годов ХХ столетия (Максимук, Проняев, 1986; Банников и др., 1961; жирнов, 1982; жирнов и др., 1998; Фадеев, Слудский, 1982). Совместный анализ эмбриональной плодовитости и размерных характеристик самок показал, что именно крупные самки, попадающие под действие минус-вектора отбора промысловой элиминации, имеют наибольшую частоту встречаемости двойневых беременностей (табл. 18).
Из экологических характеристик, изменившихся в конце 80-х годов прошлого столетия и сохраняющихся по настоящее время, отметим прекращение сезонных перекочёвок (миграций) калмыцкой популяции. Места отёла, зимовок и летовок животных совместились. Популяция практически круглый год проводит в местах прежних зимовок на юге ареала (Букреева, 2000). Следует также отметить, что средний возраст животных сократился и составил для самцов – 2,0 года, самок – 2,7 года, что примерно в 2 раза меньше, чем в конце 50-х годов (жирнов, Проняев и др., 1998).
Популяция потеряла промысловое значение, последний промысел проводился в 1996 году после длительного, практически 10-летнего перерыва. Численность современной группировки колеблется в пределах 10–20 тысяч животных.
Второй пример, касающийся копытных животных, относится к популяции северного оленя (Rangifer tarandus), обитающего на о. Врангеля (Давыдов, 1997). Особенностью опромышления, позволившей выявить направление вектора промысловой элиминации, является следующее. Для добычи животных в районе пос. Ушаковский был оборудован кораль, в который после установления устойчивых холодов с помощью мотонарт загонялись олени5. К моменту сбора материала (1991 г.) с помощью кораля в течение нескольких лет проводилась добыча животных, обитающих в восточной части острова, наиболее доступной для проведения этих работ. В 1991 году впервые произведен загон животных в кораль из западной части острова, что позволило сравнить характеристики опромышляемой в течение нескольких лет группировки с неопромышляемой.
Во время добычи обследовался ряд количественных признаков экстерьера. Результаты показали, что практически для всех проанализированных признаков их средние значения у животных из западной части острова, ранее не облавливаемой, были выше.
Втаблице 19 в качестве примера приведены значения признака «масса туши». Принимая во внимание, что размеры острова невелики, можно с высокой степенью
достоверности предполагать, что западная и восточная части популяции обитают в отно-
Таблица 18
Взаимосвязь размеров самок и их эмбриональной плодовитости
Градации признака |
Частота двойневых |
косая длина туловища, см |
беременностей, % |
73–75 |
0 |
76–78 |
0 |
79–81 |
0.05 |
82–84 |
0.05 |
85–87 |
0.13 |
88–90 |
0.37 |
91–93 |
0.67 |
5 Принципы управления пространственным поведением стада аналогичны тем, что описаны для сайгака (Максимук, Проняев, 1983).
309