

40 |
Запоры у детей |
го микробиоценоза. С биохимической точки зрения под пребиотиками понимают обычно углеводные компоненты питания, особо необходимые для жизнедеятельности нормальной кишечной микрофлоры. Залогом поступления к бактериям необходимых нутриентов является рациональное питание макроорганизма и нормальное течение пищеварительных процессов.
Хотя моносахариды могут легко утилизироваться микроорганизмами толстой кишки, к пребиотикам их не относят, т.к. они должны полностью всасываться в тонкой кишке. К пребиотикам относят некоторые дисахариды, олигосахариды, полисахариды и достаточно гетерогенную группу соединений, которую обозначили как пищевые волокна. В женском молоке из пребиотиков присутствует лактоза и олигосахариды.
Лактоза (молочный сахар) представляет собой дисахарид, состоящий из галактозы и глюкозы. В норме лактоза расщепляется лактазой тонкой кишки до мономеров, которые практически полностью всасываются в тонкой кишке. Лишь незначительное количество нерасщепленной лактозы у детей первых месяцев жизни попадает в толстую кишку, где утилизируется микрофлорой, обеспечивая ее становление.
Олигосахариды представляют собой линейные полимеры глюкозы и других моносахаров с общей длиной цепи не более десяти. По химической структуре выделяют галакто-, фрукто-, фукозилолигосахариды и др. Концентрация олигосахаридов в женском молоке относительно невелика, не более 12–14 г/л, однако их пребиотический эффект весьма значителен. Именно олигосахариды сегодня рассматриваются как основные пребиотики женского молока, обеспечивающие как становление нормальной микрофлоры кишечника ребенка, так и поддержание в дальнейшем.
Полисахариды представляют собой длинноцепочечные углеводы, в основном растительного происхождения. Инулин, содержащий фруктозу, в больших количествах присутствует в артишоках, клубнях и корнях георгинов и одуванчиков. Утилизируется бифидо- и лактобактериями, способствует их росту. Помимо этого, инулин повышает всасывание кальция
ивлияет на метаболизм липидов, снижая риск развития атеросклероза. Пищевые волокна – большая гетерогенная группа полисахаридов, наи-
более известными из которых являются целлюлоза и гемицеллюлоза. Целлюлоза – неразветвленный полимер глюкозы, а гемицеллюлоза – полимер глюкозы, арабинозы, глюкуроновой кислоты и ее метилового эфира. Помимо субстрата для питания лакто- и бифидофлоры и таким образом поставщика короткоцепочечных жирных кислот для колоноцитов, пищевые волокна оказывают и другие важные эффекты. Они обладают высокой адсорбционной способностью и удерживают воду, что приводит к повышению осмотического давления в полости кишки, увеличению объе-
Анатомо-физиологические особенности толстой кишки и акта дефекации у детей |
41 |
ма фекалий, ускорения пассажа по кишечнику, что обуславливает слабительный эффект.
В средних количествах (1–1,9 г/100 г продукта) пищевые волокна содержатся в моркови, сладком перце, петрушке (в корне и зелени), редьке, репе, тыкве, дыне, черносливе, цитрусовых, бруснике, фасоли, гречневой, и перловой крупе, «Геркулес», ржаном хлебе.
Высокое содержание (2–3 г/100 г продукта) пищевых волокон характерно для чеснока, клюквы, красной и черной смородины, черноплодной рябины, ежевики, овсяной крупы, хлебе из белково-отрубной муки.
Наибольшее же их количество (более 3 г/100 г) содержится в укропе, кураге, клубнике, малине, чае (4,5 г/100 г), овсяной муке (7,7 г/100 г), пшеничных отрубях (8,2 г/100 г), сушеном шиповнике (10 г/100 г), жареном кофе в зернах (12,8 г/100 г), овсяных отрубях (14 г/100 г). Пищевые волокна не содержатся в рафинированных продуктах.
Пребиотики важны не только для питания микрофлоры, но также для благополучия ЖКТ и всего организма в целом. В то же время, в современных условиях отмечается дефицит пребиотиков в питании во всех возрастных группах. В частности, взрослый человек должен съедать в сутки примерно 20–35 г пищевых волокон, тогда как в реальных условиях европейский житель потребляет не более 13 г в сутки. Уменьшение доли естественного вскармливания у детей первого года жизни приводит к недостатку пребиотиков, содержащихся в женском молоке. Эта нехватка пребиотиков в питании может объяснять, как увеличение частоты функциональных запоров, так и увеличение частоты рака толстой кишки.
Таким образом, пребиотики обеспечивают благополучие микрофлоры толстойкишки,еездоровьеиявляютсянеобходимымфакторомздоровьячеловека
всвязисихсущественнымиметаболическимиэффектами.Преодолениедефицитапребиотиковвсовременныхусловияхсвязанособеспечениемрациональногопитаниячеловекавовсехвозрастах:отноворожденногодопреклонного.
Свои энергетические потребности микроорганизмы толстой кишки обеспечивают за счет анаэробного субстратного фосфорилирования, в ходе которого происходит образование преимущественно молочная кислота (молочное брожение, характерное для лактобактерий, стрептококков толстой кишки) или молочная кислота в сочетании с другими метаболитами (в т.ч. уксусной кислотой, присущее бифидобактериям). Спиртовое брожение, ведущее к образованию углекислого газа и этанола, является побочным метаболическим путем у некоторых представителей Lactobacillus и Clostridium. Отдельные виды энтеробактерий (E.coli) и клостридий получают энергию
врезультате муравьинокислого, пропионовокислого, маслянокислого, ацетонобутилового или гомоацетатного видов брожения.
42 |
Запоры у детей |
Врезультатемикробногометаболизмавтолстойкишкеобразуютсямолочная кислота, короткоцепочечные жирные кислоты, углекислый газ, водород, вода.Углекислыйгазвбольшойстепенипреобразуетсявацетат,водородвсасывается и выводится через легкие, а органические кислоты утилизируются макроорганизмом, причем значение их для человека трудно переоценить.
Говоря о жирных кислотах, наибольшее значение имеют уксусная (обозначается как С2, т.е. содержит 2 углеродных атома), пропионовая (С3), масляная / изомасляная (С4), валериановая / изовалериановая (С5), капроновая / изокапроновая (С6). Перечисленные жирные кислоты в медицинской литературе обозначаются как короткоцепочечные, хотя с биохимической точки зрения таковыми являются только три первых (т.е. С2-С4). Нормальная микрофлора толстой кишки перерабатывая непереваренные в тонкой кишке углеводы производит перечисленные кислоты с минимальным количеством их изоформ. В то же время, при нарушении микробиоценоза и увеличении доли протеолитической микрофлоры указанные жирные кислоты начинают синтезироваться из белков преимущественно в виде изоформ, что отрицательным образом сказывается на состоянии толстой кишки, с одной стороны, и может быть диагностическим маркером, с другой.
Концентрация короткоцепочечных жирных кислот (КЦЖК) в толстой кишке максимальная в проксимальных отделах, т.е. там, где идет наиболее интенсивный их синтез, и снижается к дистальным отделам. В среднем концентрация КЦЖК в толстой кишке взрослого человека составляет 70–140 ммоль/л в проксимальных отделах толстой кишки и 20–70 ммоль/л – в дистальных [15]. У новорожденного ребенка КЦЖК в толстой кишке в виду отсутствия микрофлоры практически отсутствуют. Их количество у здорового ребенка резко нарастает к 20–30 дню жизни до 70–80 ммоль/л, что совпадает с периодом нормального становления кишечной микрофлоры и далее достигает «взрослых» значений к 2-м годам жизни [16].
Соотношение концентраций С2:С3:С4 в толстой кишке взрослого человека составляет примерно 57:21:22%. Остальные КЦЖК присутствуют в незначительных количествах. Ацетат является преобладающим во всех возрастных группах, однако у детей первых месяцев жизни, находящихся на естественном вскармливании, концентрация бутирата и пропионата очень невелики, а основную часть метаболитов микробной микрофлоры составляют ацетат и лактат. При искусственном вскармливании доля лактата уменьшается, а бутирата и пропионата увеличивается, что объясняется изменением состава микрофлоры и может иметь отрицательные последствия для состояния кишечника [17].
КЦЖК, образованные в результате микробного метаболизма, имеют важное значение как для толстой кишки, так и для макроорганизма в целом.
Анатомо-физиологические особенности толстой кишки и акта дефекации у детей |
43 |
Синтез КЦЖК является важным фактором колонизационной резистентности, обеспечивающим стабильность состава кишечной микрофлоры, одним, но не единственным, механизмом обеспечения которой является поддержание оптимальных значений рН в просвете толстой кишки. Повышение концентрации КЦЖК сочетается со снижением осмотического давления в толстой кишке в связи с расщеплением полисахаридов [15, 18–20].
Регуляциякишечноготранзитаорганическимикислотамисвязананетолько с изменением осмотического давления в результате их синтеза. В проксимальных отделах толстой кишки КЦЖК стимулируют рецепторы L-клеток, которые вырабатывают регуляторный пептид PYY, который, в свою очередь, замедляет моторику как толстой, так и тонкой кишки. Установлено, что вы-
работка PYY лежит в основе «илеоцекального тормоза», замедляющего ки-
шечную моторику при попадании в толстую кишку недопереваренных жиров [21]. В дистальных отделах толстой кишки эффект КЦЖК противоположный. Они стимулируют рецепторы Ecl-клеток, вырабатывающих гистамин, который действуя на 5-HT4-рецепторы афферентных волокон блуждающего нерва инициирует рефлекторное ускорение моторики [22].
Большая часть КЦЖК, образовавшихся в толстой кишке, всасывается. Обычно, с калом выводится не более 5% от их общего количества. Всасывание КЦЖК происходит при участии активных транспортных систем колоноцитов и наиболее хорошо изучено в отношении бутирата. Установлено, что бутират поступает в колоноцит в обмен на гидрокарбонатные ионы. Часть всосавшегося бутирата поступает опять в просвет кишки в обмен на ионы хлора, однако значительная часть его остается в колоноците и утилизируется им. Кроме того, всасывание бутирата тесно связано с всасыванием натрия: блокированиевсасываниебутиратаблокируетвсасываниенатрияинаоборот. Это взаимодействие имеет особое значение, т.к. поступление натрия в колоноцит определяет всасывание воды. Кроме того, КЦЖК определяют всасывание кальция и магния. Таким образом, эффективность всасывания КЦЖК имеет значение не только для поддержания водно-электролитного равновесия и минерального обмена в организме, но также для регуляции моторики толстой кишки, проявляя свой антидиарейный эффект [23, 24].
Наконец, важнейшей функцией микрофлоры в связи с метаболизмом пребиотиков в КЦЖК является обеспечение колоноцита энергией, который для энергетических целей не менее чем на 70% потребляет бутират и значительнострадаетприегонедостатке.Показано,чтоКЦЖКявляютсярегуляторами апоптоза и обладают антиканцерогенным эффектом, снижая пролиферацию клеток эпителия толстой кишки, но повышая ее их дифференцировку [25].
Поступившиевколоноцитацетатипропионатвосновномвыводятсявкровоток. При этом на уровне толстой кишки они участвуют в регуляции ее кро-
44 |
Запоры у детей |
вотока,повышаяееитемсамымобладаютантиишемическимэффектом.Концентрация КЦЖК (в основном, ацетат и пропионат) в воротной вене составляетвсреднем375±70ммоль/л,втовремякаквоттекающейотпеченикрови онаснижаетсядо148±42ммоль/л,авпериферическойкрови–79±22ммоль/л. Таким образом, печень задерживает примерно половину поступивших через колоноцит КЦЖК, а периферические ткани элиминируют еще одну четверть их. Большая часть ацетата и пропионата в тканях идет на синтез глюкозы и небольшая часть (не более 10%) на энергетические нужды [15].
Таким образом, метаболическая активность микрофлоры толстой кишки имеет существенное значение для благополучия ЖКТ и всего организма в целом, в частности, обеспечения нормального кишечного транзита.
Физиология дефекации
Дефекация является сложным скоординированным рефлекторным актом, в котором принимают участие кора головного мозга, ВНС, рецепторы прямой кишки, мускулатура брюшного пресса и толстой кишки. На первом году жизни ребенка дефекация протекает по типу безусловного рефлекса, в то время как со второго года жизни начинают формироваться условно-рефлекторные связи с временем суток (утро, вечер), местом (туалетная комната; детский горшок), звуками и мн.др.
Пропульсивные сокращения толстой кишки обычно возникают утром, вскоре после пробуждения и могут усиливаться под действием различных факторов, включая прием пищи или питье (желудочно-кишечный рефлекс), что проявляется позывами на дефекацию утром после еды. Во время сна в норме моторика толстой кишки ослабевает, что предотвращает ночное недержание кала. Каловые массы, поступая в прямую кишку, растягивают ее стенки, что вызывает появление осознанного позыва к дефекации. В том случае, если момент для опорожнения кишечника неподходящий, прямая кишка адаптируется для временного хранения каловых масс, а позывы и перистальтика угасают. У взрослых лиц период транзита содержимого по толстой кишке в норме составляет 20–72 ч [26–28].
Важным феноменом, обеспечивающим контроль над эвакуацией кишечного содержимого, является так называемое «держание» или «удержание». Осуществляется держание функциональной системой, состоящей из промежностного отдела прямой кишки с внутренним сфинктером, наружного сфинктера и мышцы, поднимающей задний проход, изгибы прямой кишки (например, аноректальный угол), поперечные складки и геморроидальное сплетение в анальном канале, образующие пассивные барьеры [29].
Различают два типа держания: кишечное и анальное, представляющие собой последовательные этапы единого механизма опорожнения кишечника.
Анатомо-физиологические особенности толстой кишки и акта дефекации у детей |
45 |
Кишечное держание обеспечивает продвижение содержимого по кишке в течение более-менее длительного периода времени, чему способствуют форма толстой кишки в виде обода, гаустрация, координированные антиперистальтические движения, ректосигмоидный изгиб, сдерживающий продвижение кишечного содержимого к прямой кишке. Данный этап держания является неосознанным.
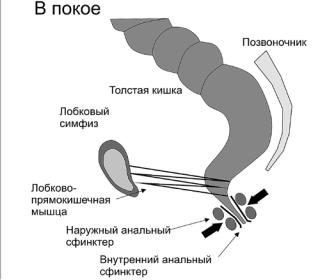
Анальное держание осуществляется за счет рефлекторного и произвольного сокращения запирательного аппарата прямой кишки: внутреннего и наружного сфинктеров. Внутренний сфинктер заднего прохода не подчиняется сознательному контролю, играя роль постоянного пассивного замыкателя. Он состоит из гладких мышц, на 70% обеспечивая тоническое смыкание стенок заднепроходного канала в покое. Активное сокращение замыкательного аппарата достигается при помощи наружного сфинктера.
Наружный сфинктер заднего прохода подчиняется волевому контролю, состоит из поперечнополосатых мышц и на 30% обеспечивает тонус заднего прохода в покое. Сокращение лобково-прямокишечной мышцы в покое поддерживает аноректальный угол, а при натуживании уменьшает его. Растяжение прямой кишки газами или каловыми массами приводит к рефлекторному расслаблению внутреннего сфинктера заднего прохода, что известно как ректоанальный тормозной рефлекс. Этот рефлекс регулируется энтеральной нервной системой без участия периферической и центральной нервных систем.
Действие наружного сфинктера реализуется с помощью трех петель: верхней, средней и нижней. Верхнюю петлю образует т.н. аноректальное кольцо, фиксирующееся к внутренней нижней поверхности лонной кости, среднюю – поверхностная порция наружного сфинктера, прикрепляющаяся к копчику, а нижнюю – подкожная порция этой мышцы, прикрепленная к перианальной коже в области срединного шва промежности. Верхняя и нижняя петли, иннервируемые геморроидальными ветвями полового нерва, перетягивают заднюю анальную стенку кпереди, в то время как средняя петля, иннервируемая четвертым крестцовым нервом, тянет переднюю анальную стенку кзади. Названные петли являются по существу отдельными сфинктерами и, взаимодействуя, обеспечивают полное держание кишечного содержимого [28] (рис. 4).
Перистальтику и функцию запирательного аппарата прямой кишки регулируют главным образам три нервных центра: 1) ганглии стенки прямой кишки и нижнее подчревное сплетение (plexus hypogastricus inf.); 2) спинномозговой центр на уровне SII-IV; 3) кора головного мозга. Регуляция функции осуществляется тремя рефлекторными дугами. Первая дуга регулирует расслабление внутреннего сфинктера и перистальтические движе-
46 |
Запоры у детей |
Рис. 4. Строение внутреннего (ВСЗП) и наружного сфинктеров заднего прохода (НСЗП) в покое и лобково-прямокишечной мышцы [30]
ния ректоанального сегмента через интрамуральный путь и тазовые сплетения, в частности подчревные. Нейроны спинномозгового центра координируют перистальтические движения и контролируют непроизвольное действие сфинктерного аппарата.
Вторую дугу образуют афферентный путь, проводящий импульсы от ректальных рецепторов к нейронам спинномозгового центра, и эфферентный путь, проводящий импульсы от спинномозгового центра к лонно-прямокишечной мышце и наружному сфинктеру. Этот рефлекс возникает при повышении давления в прямой кишке или при растяжении ее стенки и вызывает сокращение лонно-прямокишечной мышцы и наружного сфинктера, таким образом обеспечивая держание кала.
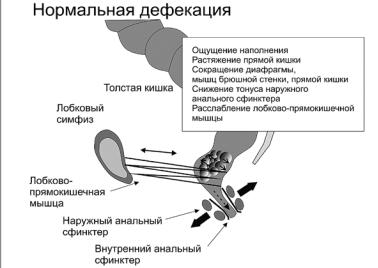
Через третью дугу кора головного мозга управляет функцией наружного сфинктера и мышц, поднимающих задний проход. Афферентные импульсы от барорецепторов леваторной мышцы через половой нерв и спинной мозг достигают коры головного мозга, и оттуда эфферентные импульсы через крестцовый спинномозговой центр возвращаются к наружному сфинктеру и леваторной мышце. По этой дуге чувство позыва к дефекации передается к коре головного мозга, и при помощи произвольного сокращения или расслабления наружного сфинктера и леваторной мышцы обеспечиваются условия для держания или дефекации (рис. 5).
Анатомо-физиологические особенности толстой кишки и акта дефекации у детей |
47 |
Рис. 5. При дефекации сфинктеры заднего прохода и лобково-прямокишечная мышца расслабляются, а мышцы брюшной стенки и прямой кишки сокращаются, что способствует выталкиванию кала из прямой кишки [30]
Если держание кишечного содержимого – относительно пассивный процесс и может осуществляться непроизвольно за счет самоуправления ректального и спинномозгового центров, то физиологическая дефекация является активным процессом и осуществляется под контролем сознания. Акт дефекации состоит из двух фаз.
В первой, непроизвольной, кишечное содержимое поступает пропульсивной волной в прямую кишку, при растяжении которой расслабляется внутренний сфинктер, давление на его уровне становится ниже, чем в прямой кишке, и кишечное содержимое продвигается дистальнее до входа в анальный канал, вступая в контакт с чувствительной зоной его слизистой оболочки. Вскоре тонус сфинктера восстанавливается, а кишечное содержимое остается выше анального канала. В первой фазе держание обеспечивается за счет рефлекторного сокращения (по второй дуге) наружного сфинктера и лонно-прямокишечной мышцы. Последняя, перетягивая кишку кзади, закрывая вход в анальный канал. В это время перистальтика усиливается, кишечное содержимое поступает в ампулу, последняя еще больше растягивается; раздражаются не только рецепторы ампулы прямой кишки, но и леваторные мышцы. Импульсы достигают коры головного мозга по третьей рефлекторной дуге, возникает позыв к дефекации и наступает следующая, произвольная, фаза. Если условия для дефекации социально
48 |
Запоры у детей |
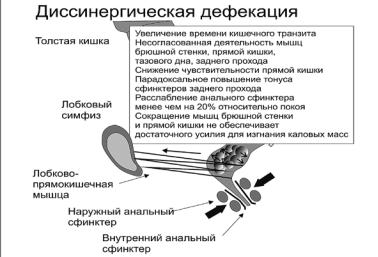
Рис. 6. При дискинетическом запоре сокращение мышц, необходимое для продвижения кала к заднепроходному каналу, и расслабление сфинктеров заднего прохода и лобковопрямокишечной мышцы могут нарушаться, что ведет к несогласованной работе мышц при попытке дефекации (диссинергии) и задержке кала [30]
приемлемы, то при анализе чувства позыва кора головного мозга подает команду совершить акт дефекации (рис. 6).
Наружный сфинктер произвольно расслабляется, увеличивается аноректальный угол, внутренний сфинктер полностью раскрыт и вследствие повышения внутрибрюшного давления кал эвакуируется наружу [1].
Наружный сфинктер произвольно расслабляется, увеличивается аноректальный угол, внутренний сфинктер полностью раскрыт и вследствие повышения внутрибрюшного давления кал эвакуируется наружу [1].
Ключевые рецепторы для раскрытия анального канала локализуются в мышце, поднимающей задний проход. Эти рецепторы в основном контролируют непроизвольное и произвольное открытие и закрытие анального канала, их раздражение вызывает чувство позыва к дефекации. Сенсорные механизмы отвечают за восприятие содержимого прямой кишки, такого как газы или стул. В отсутствие условий для дефекации позывы на нее подавляются произвольным сокращением наружного сфинктера. Расслабление прямой кишки также подавляет позывы на дефекацию, что способствует дальнейшему накоплению кала. При наличии подходящих условий для дефекации человек принимает положение сидя (или на корточках), задерживает дыхание, напрягает диафрагму, мышцы передней брюшной стенки и прямой кишки и одновременно расслабляет наружный сфинктер
Анатомо-физиологические особенности толстой кишки и акта дефекации у детей |
49 |
и лобково-прямокишечную мышцу [30]. Все это приводит к открытию заднего прохода и выделению кала (рис. 4).
Таким образом, нормальное функционирование прямой кишки и заднего прохода включает в себя сенсорное восприятие и координацию физиологических процессов.
Список литературы
1.Ленюшкин А.И. Детская колопроктология. М., 1990. 352 с.
2.Andrews Ch.N., Storr M. Pathophysiology of chronic constipation. Can J Gastroenterol. 2011; 25 Supple В: 17B–23B.
3.Lynch AC, Anthony A, Dobbs BR, Frizelle FA. Anorectal physiology following spinal cord injury. Spinal Cord.- 2000. 38:573-580.
4.Aaronson MJ, Freed MM, Burako R. Colonic myoelectric activity in persons with spinal cord injury. Dig Dis Sci. 1985. 30:295-300.
5.Rostad H. Colonic motility in the cat. II. Extrinsic nervous control. Acta Physiol Scand.- 1973. 89: 91-103.
6.Lysy J, Karmeli F, Sestieri M, Yatzkan Y, Goldin E. Decreased substance P content in the rectal mucosa of diabetics with diarrhoea and constipation. Metabolism.- 1997. 46: 730-734.
7.Simrén M, Barbara G, Flint HJ, Spiegel BM, Spiller RC, Vanner S, Verdu EF, Whorwell PJ, Zoetendal EG. Intestinal microbiota in functional bowel disorders: a Rome foundation report. Gut 2013; 62: 159-176.
8.Dethlefsen L, McFall-Ngai M, Relman DA. An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 2007; 449: 811-818.
9.Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, Mende DR, Li J, Xu J, Li S, Li D, Cao J, Wang B, Liang H, Zheng H, Xie Y, Tap J, Lepage P, Bertalan M, Batto JM, Hansen T, Le Paslier D, Linneberg A, Nielsen HB, Pelletier E, Renault P, Sicheritz-Ponten T, Turner K, Zhu H, Yu C, Li S, Jian M, Zhou Y, Li Y, Zhang X, Li S, Qin N, Yang H, Wang J, Brunak S, Doré J, Guarner F, Kristiansen K, Pedersen O, Parkhill J, Weissenbach J, Bork P, Ehrlich SD, Wang J. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010; 464: 59-65.
10.Faith JJ, Guruge JL, Charbonneau M, Subramanian S, Seedorf H, Goodman AL, Clemente JC, Knight R, Heath AC, Leibel RL, Rosenbaum M, Gordon JI. The longterm stability of the human gut microbiota. Science 2013; 341: 1237439.
11.Koren O, Knights D, Gonzalez A, Waldron L, Segata N, Knight R, Huttenhower C, Ley RE. A guide to enterotypes across the human body: meta-analysis of microbial community structures in human microbiome datasets. PLoS Comput Biol 2013; 9: e1002863.
12.ArumugamM,RaesJ,PelletierE,LePaslierD,YamadaT,MendeDR,FernandesGR,Tap J, Bruls T, Batto JM, Bertalan M, Borruel N, Casellas F, Fernandez L, Gautier L, Hansen T, Hattori M, Hayashi T, Kleerebezem M, Kurokawa K, Leclerc M, Levenez F, Manichanh C, Nielsen HB, Nielsen T, Pons N, Poulain J, Qin J, Sicheritz-Ponten T, Tims S, Torrents D, Ugarte E, Zoetendal EG, Wang J, Guarner F, Pedersen O, de Vos WM, Brunak S, Doré J, Antolín M, Artiguenave F, Blottiere HM, Almeida M, Brechot C, Cara C, Chervaux C, Cultrone A, Delorme C, Denariaz G, Dervyn R, Foerstner KU, Friss C, van de Guchte M, Guedon E, Haimet F, Huber W, van Hylckama-Vlieg J, Jamet A, Juste C, Kaci G, Knol J,
Lakhdari O, Layec S, Le Roux K, Maguin E, MérieuxA, Melo Minardi R, M’rini C, Muller J,