

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdf11.4. Болевая рецепция (ноцицепция)
Ноцицептивные волокна не имеют специализированных оконча- ний, а представляют собой свободные нервные окончания в коже и многих других частях организма. Ноцицепция возникает из сочетания прямых и непрямых воздействий на периферические сенсоры. Сильное нагревание вызывает открытие неспецифических катионных каналов в окончаниях С-волокон. Входящие Na+- è Ñà2+-токи деполяризуют волокно, что приводит к генерации нервных импульсом в клетке. Кислоты (протоны) также напрямую могут открывать катионные каналы. Кроме того, деполяризацию ноцицепторов могут вызывать и механически воздействия.
При повреждениях тканей выделяются различные химические активаторы, например АТФ. На мембранах нейронов ганглиев задних корешков спинного мозга, образующих С-волокна, найдены пуриновые рецепторы P2X3. АТФ, воздействуя на эти лиганд-активируемые рецепторы, открывает неспецифические катионные каналы, что приводит к возбуждению ноцицепторов.
Выводы
•Внешние сигналы разной физической природы воздействуют на специализированные сенсорные рецепторы, в результате чего в них возникают электрические потенциалы. По механизму активации сенсорные рецепторы подразделяют на механо-, хемо-, фото- и терморецепторы.
•Механочувствительные ионные каналы обнаружены повсеместно: в эндотелиальных клетках кровеносных сосудов, в барорецепторах синусов, в рецепторах прикосновения и давления в коже, в мышечных рецепторах и в механочувствительных волосковых клетках внутреннего уха позвоночных и органа боковой линии у рыб.
•Хеморецепция обеспечивает чувствительность организмов к молекулярному составу окружающей среды. В отличие от механорецепторов, молекулярный механизм трансдукции химического сигнала хеморецепторами является общим для многих из них и основан на преобразовании сигнала с участием G-белков.
•В коже млекопитающих на изменения температуры реагируют свободные нервные окончания. Аδ- (холодовые) и С-афференты (тепловые) от кожных терморецепторов оканчиваются в дорзальных рогах спинного мозга.
290
•Болевая рецепция (ноцицепция) возникает из сочетания прямых
èнепрямых воздействий на периферические сенсоры.
Цитированная литература
Рекомендуемая
1.Ашмарин И. П. Биохимия мозга. СПб.: Изд-во С.-Петербургского ун-та,
1999.
2.Николс Дж. Г., Мартин А. Р., Валлас Б. Дж., Фукс П. А. От нейрона к мозгу. М.: Едиториал УРСС, 2003.
3.Смит К. Биология сенсорных систем. М.: Бином. Лаборатория знаний,
2005
4.Шульговский В. В. Физиология центральной нервной системы. М.: МГУ,
1997.
Дополнительная
1.Гранит Г. Основы регуляции движений. М.: Мир, 1973.
2.Катц Б. Нерв, мышца, синапс. М.: Мир, 1968.
3.Zigmond M. J., Bloom F. E., Landis S. C., Roberts J. LSquire., L. R. Fundamental Neuroscience. Academic Press, San Diego, London, 1999.
291
Глава 12. РЕЦЕПЦИЯ В СЕНСОРНЫХ СИСТЕМАХ: ФОТОРЕЦЕПТОРЫ
Рассмотрены общая морфология фоторецепторов сетчатки позвоночных животных, механизмы генерации светового ответа и выделения медиатора фоторецепторами. Приводятся современные данные о молекулярных механизмах фототрансдукции, о роли потенциал-зависимых ионных каналов, рецепторов ГАМК, переносчиков и рецепторов глутамата в модуляции выделения медиатора синаптическими терминалями фоторецепторных клеток.
12.1.Морфология фоторецепторов сетчатки позвоночных
12.1.1.Общая характеристика
Óвсех позвоночных животных, включая человека, сетчатка содержит два типа фоторецепторов, различающихся морфологически и функционально. Колбочки — относительно крупные клетки конической формы, преобладают у животных, ведущих дневной образ жизни. Палочки — более тонкие и вытянутые клетки, обладающие значительно большей чувствительностью к свету, чем колбочки, доминируют в сет- чатке животных с ночным образом жизни (Cajal, 1892; Polyak, 1941; Dowling, Boycott, 1966; Shkolnik-Yarros, 1971 è äð.).
Колбочки сетчатки млекопитающих относятся к одному морфологическому типу — одиночные колбочки, тогда как в сетчатке низших позвоночных представлены не только одиночные, но и двойные колбочки, состоящие из основного и дополнительного члеников (Cajal, 1892; Marc, Sperling, 1976).
Óбольшинства млекопитающих выявлены палочки и один—два типа колбочек, различающихся по спектральной чувствительности. Только в сетчатке приматов и человека представлены три типа колбо- чек, чувствительных в синей (λ = 445 нм), зеленой (λ = 535 нм) и красной (λ = 570 нм) областях спектра (Gouras, 1984). Колбочки, чувствительные к коротковолновому свету, обладают некоторыми морфологическими особенностями, отличающими их от остальных фоторецепторов (Ahnelt e.a., 1990).
Кроме того, в отличие от зелено- и красночувствительных колбо- чек, распределенных по сетчатке практически одинаково и равномерно, колбочки, чувствительные к синему свету, относительно малочис-
292
ленны и составляют 8% , 3-5% и 15% от общего числа колбочек на периферии сетчатки, в центре и на периферии фовеа, соответственно (Ahnelt e.a., 1987).
Спектральные характеристики колбочек у домлекопитающих позвоночных отличаются значительно большим разнообразием, а рыбы семейства карповых являются своего рода «приматами среди рыб». В сетчатке этих животных наряду с палочками представлены три типа колбочек, содержащих зрительные пигменты с максимумом чувствительности в синей, зеленой либо красной областях спектра (λ=445 нм, λ=535 нм и λ=570 нм).
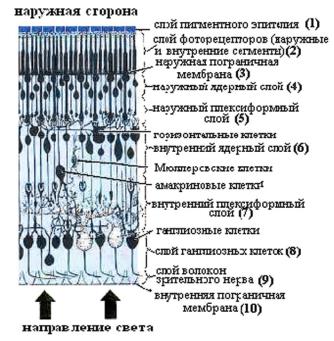
Ðèñ. 12.1. Основные клеточные элементы сетчатки (по Polyak, 1941).
Пигменты с различными спектральными характеристиками содержатся, как правило, в разных фоторецепторах. До недавнего времени исключением считались двойные колбочки низших позвоночных, имеющие неодинаковые спектральные характеристики пигмента дополнительной и основной колбочки (Liebman, Entine, 1968; Marc, Sperling,
293
1976). Однако оказалось, что в сетчатке мыши наряду с сине- и зелено- чувствительными колбочками, содержащими пигменты, чувствительные к свету, с длиной волны 360 нм и, соответственно, 508 нм, есть колбочки, содержащие оба пигмента. (Applebury e.a., 2000).
12.1.2. Распределение фоторецепторов по сетчатке.
Фоторецепторы обоих типов ориентированы перпендикулярно к поверхности сетчатки и образуют наружные слои, наиболее удаленные (дистальные) от света, падающего на сетчатку (рис. 12.1).
Первый наружный слой сетчатки образуют клетки пигментного эпителия (1), примыкающего к сосудистой оболочке глаза. Второй слой (2) сформирован наружными и внутренними сегментами фоторецепторов, а четвертый — наружный ядерный слой (4) — образуют тела фоторецепторов, содержащие клеточные ядра. Второй и четвертый слои разделены так называемой наружной плазматической мембраной (слой 3), образованной горизонтальными отростками глиальных (Мюллеровских) клеток. Аксоны фоторецепторов, отходящие от сомы фоторецепторных клеток, и контактирующие с ними отростки биполярных и горизонтальных клеток образуют наружный сетевидный (плексиформный) слой (5).
Распределение палочек и колбочек по сетчатке неодинаково. У млекопитающих палочки, составляющие подавляющее большинство фоторецепторов на периферии сетчатки, весьма малочисленны в центральной зоне (area centralis) либо совсем отсутствуют в области фовеа (центральной ямки, центра сетчатки) сетчатки приматов. Количество колбочек, наоборот, возрастает в центральной зоне сетчатки и убывает на периферии. В фовеа приматов и человека численность колбочек настолько велика, что расстояние между центрами фоторецепторов составляет 2–3 мкм, а сами колбочки становятся тоньше, чем палочки, и образуют мозаику в форме плотно упакованных гексагональных образований. Большая плотность фоторецепторов обеспечивает высокую разрешающую способность (1 угловая минута) этой области сетчатки (Wassle, Boycott, 1991).
У низших позвоночных так же, как у млекопитающих, фоторецепторы определенным образом упорядочены и образуют своеобразную мозаику, обнаруживающуюся у всех животных начиная с рыб (Marc, Sperling, 1976; Подугольникова, Максимов, 1977). Некоторые аналоги центральной зоны и фовеа сетчатки млекопитающих и приматов также можно найти на самых ранних уровнях филогенеза. Так, многие виды рыб имеют одну (золотой карась) или две (гуппи) области сетчатки с
294
повышенной плотностью колбочек, а у некоторых подвижных морских видов (род Serranus), обитающих в прибрежной зоне, есть ямкообразное углубление, содержащее большое количество колбочек и напоминающее фовеа приматов (Гусельников, Логинов, 1976).
У рыб преобладание того или иного типа фоторецепторов отчетливо кореллирует с условиями освещения в среде обитания. Так, исследование сетчатки рыб в онтогенезе показало, что в процессе развития чисто колбочковая сетчатка личинок, обитающих на мелководье, сменяется у взрослых особей сетчаткой смешанного типа с преобладанием палочек (Школьник-Ярос, Бабурина, 1982).
12.1.3. Строение фоторецептора
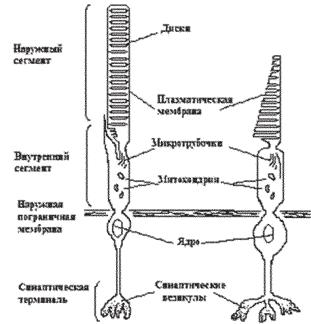
Фоторецептор (как палочка, так и колбочка) состоит из связанных между собой наружного и внутреннего сегментов, тела клетки и синаптического окончания («аксона») фоторецептора (рис. 12.2).
À. Палочка |
Á. Колбочка |
Ðèñ. 12.2. Строение фоторецепторов. Схематическое изображение палочки (А), колбочки (Б).
295
Наружный сегмент фоторецептора имеет колбочковую или палоч- ковую форму и заполнен плотно упакованными (расстояние между дисками около 28 нм) мембранными дисками, содержащими зрительный пигмент и другие компоненты фототрансдукции. У всех позвоноч- ных животных плотность молекул зрительного пигмента в мембране диска очень велика — около 25000 молекул на 1 квадратный микрон, что соответствует концентрации пигмента около 3,5 мМ (Harosi, 1975). Высокая концентрация пигмента в дисках и плотная упаковка самих дисков обеспечивают большую вероятность захвата фотона.
Процесс формирования дисков, заполняющих наружный сегмент, у палочек и колбочек неодинаков. Складки — впячивания клеточной мембраны палочек — отделяются в виде свободных дисков от наружной мембраны, сохраняя с нею связь только у начала наружного сегмента. Таким образом, диски со встроенными молекулами зрительного пигмента оказываются у палочек изолированными от внеклеточного пространства, тогда как диски колбочек имеют непосредственный контакт с экстраклеточной средой, обеспечивающий, видимо, более быстрое проникновение веществ как из клетки, так и в клетку.
Внутренний сегмент фоторецептора содержит эндоплазматический ретикулум, аппарат Гольджи и множество митохондрий, расположенных в области, прилегающей к наружному сегменту, и обеспечивающих, видимо, энергетические потребности фототрансдукции. Несколько узких микротрубочек соединяют наружный и внутренний сегменты фоторецептора. Полагают, что они обеспечивают транспортировку высокомолекулярных белковых молекул и участвуют в формировании новых дисков.
Стопки дисков, содержащих молекулы пигмента, постоянно обновляются (Young, 1971; Steinberg e.a., 1977). Старые диски смещаются к верхушке наружного сегмента, отрываются и поглощаются (фагоцитоз) отростками пигментного эпителия, окружающими наружные сегменты фоторецепторов. В это же время в основании наружного сегмента добавляются новые диски, возникающие в области, расположенной во внутреннем сегменте у основания ресничек (Steinberg e.a., 1980). Фагоцитоз наружных сегментов преобладает у палочек в светлое, а у колбочек — в темное время суток (Young, 1971; Steinberg e.a., 1977).
От тела клетки отходит «аксон» фоторецептора, образующий специфические синаптические терминали, контактирующие с последующими клетками. У колбочек это так называемые колбочковые ножки — большое коническое (диаметр основания до 8–10 мкм) утолще-
296
ние аксона, а у палочек — сферулы — небольшое (диаметр 3–5 мкм) округлое расширение аксона. Аксональные терминали фоторецепторных клеток содержат большое количество синаптических везикул и образуют различные типы контактов как с соседними фоторецепторами, так и со вторичными нейронами сетчатки — биполярными и горизонтальными клетками.
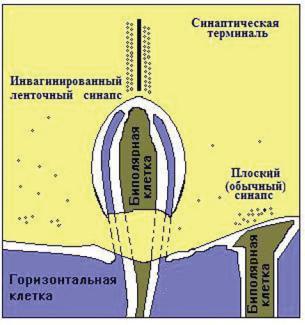
Синаптические терминали фоторецепторов образуют два типа контактов с вторичными нейронами: ленточные, или инвагинированные, и базальные, или плоские.
Ðèñ. 12.3. Схематическое трехмерное изображение инвагинированного ленточного синапса (по: Sjostrand, 1976).
В контактах первого типа (рис. 12.3) дендрит биполярной клетки расположен в инвагинации синаптической терминали фоторецептора напротив ленточного синапса, характеризующегося большим числом синаптических везикул, расположенных с обеих сторон лентообразного уплотнения (Dowling, Boycott, 1966; Kolb, 1970; Lasansky, 1971; Saito e.a., 1985). Часто постсинаптическими элементами ленточного синапса являются также два отростка горизонтальных клеток, входящие в ин-
297
вагинацию мембраны синаптической терминали фоторецептора и формирующие так называемую триаду. Центральный элемент триады образован дендритным отростком биполярной клетки, а два латеральных элемента представлены отростками горизонтальных клеток. Плоские контакты, образованные синаптическими терминалями фоторецептора, представляют собой обычные синапсы с узкой либо с широкой синаптической щелью.
Контакты обоих типов выявлены в синапсах фоторецепторов как с биполярными, так и с горизонтальными клетками. Однако у каждой отдельно взятой биполярной клетки представлен только один тип контактов — либо инвагинированные, либо плоские.
В сетчатке млекопитающих обнаружено различие между палочками и колбочками по типу контакта с биполярными клетками. У этих животных между палочками и палочковыми биполярными клетками обнаружены только контакты инвагинированного типа, тогда как между колбочками и колбочковыми биполярными клетками выявлены как инвагинированные, так и базальные контакты (Dowling, Boycott, 1966; Kolb, 1970). У низших позвоночных такой закономерности не обнаружено: в сетчатке амфибий (Lasansky, 1971) и рыб (Saito e.a., 1985) палочки и колбочки образуют с биполярными клетками как инвагинированные, так и базальные контакты.
Фоторецепторы связаны друг с другом через телодендрии — отростки, идущие от боковой или базальной части колбочковой ножки к ножкам соседних колбочек либо к сферулам палочек (Cajal, 1892; Raviola, Gilula, 1973; Raviola, Gilula, 1975; Schwartz, 1976; Ahnelt e.a., 1990).
Контакты, образуемые телодендриями с соседними фоторецепторами, представляют собой как обычные химические (Lasansky, 1971; Kolb, Jones, 1985), так и электрические синапсы (gap junction) (Raviola, Gilula, 1975; Nelson, 1977; Ahnelt e.a., 1990). Данные о зависимости межрецепторных связей через электрические синапсы от типа или спектральных характеристик фоторецепторов противоречивы. Так, было показано, что в сетчатке черепах связи между фоторецепторами упорядочены таким образом, что палочки контактируют только с палочками, а колбочки избирательно связаны с колбочками той же спектральной чувствительности (Detwiler, Hodgkin, 1979). Таким образом, фоторецепторы оказываются объединенными в два электрических синцития с различными пространственными характеристиками (Schwartz, 1976; Detwiler, Hodgkin, 1979). Однако в сетчатке всех исследованных животных: чере-
298
пах (Schwartz, 1976), кошек (Nelson, 1977) и приматов (Raviola, Gilula, 1975; Ahnelt e.a., 1990) — были обнаружены связи как между колбоч- ками с различной спектральной чувствительностью, так и между колбочками и палочками. В сетчатке приматов и человека на сферуле одной палочки выявлено 3-5 электрических контакта с телодендриями соседних колбочек, а на ножке одной зеленоили красночувствительной колбочки обнаружено не менее 10 контактов с телодендриями соседних палочек. Синечувствительные колбочки не имеют столь многочисленных электрических контактов с телодендриями соседних палочек либо колбочек и являются в некоторой степени изолированными от остальных фоторецепторов (Ahnelt e.a., 1990).
Благодаря электрическим контактам, между палочками и колбочками световая активация палочек вызывает аналогичную активацию в соседних колбочках (макаки) (Schneeweis, Schnapf, 1995) и наоборот, в палочках, электрически связанных с колбочками (10– 29% палочек), возникает ответ на освещение колбочки (саламандра) (Wu, Yang, 1988).
Полагают, что электрические контакты между палочками и колбочками обеспечивают передачу палочкового сигнала через колбочковые пути в условиях скотопической и мезопической адаптации (Demb, Pugh, 2002).
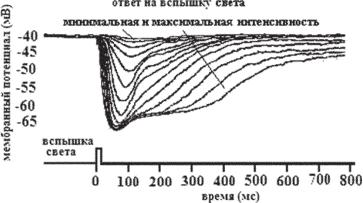
Ðèñ. 12.4. Внутриклеточная запись реакции фоторецептора (колбочки) на вспышку света переменной интенсивности. При больших интенсивностях света амплитуда ответа достигает насыщения (около -65 мВ) (по Schnapf, Baylor, 1987).
299